Johanna Pesonen CRISPR-MENETELMÄ POISTOGEENISEN SOLULINJAN VAL- MISTUKSESSA
|
|
|
- Krista Jokinen
- 8 vuotta sitten
- Katselukertoja:
Transkriptio

1 Johanna Pesonen CRISPR-MENETELMÄ POISTOGEENISEN SOLULINJAN VAL- MISTUKSESSA
2 CRISPR-MENETELMÄ POISTOGEENISEN SOLULINJAN VAL- MISTUKSESSA Johanna Pesonen Opinnäytetyö Syksy 2014 Laboratorioalan koulutusohjelma Oulun ammattikorkeakoulu
3 TIIVISTELMÄ Oulun ammattikorkeakoulu Laboratorioalan koulutusohjelma, bioteknologian sv. Tekijä: Johanna Pesonen Opinnäytetyön nimi: CRISPR-menetelmä poistogeenisen solulinjan valmistuksessa Työn ohjaajat: Sakari Kellokumpu ja Matti Nieminen Työn valmistumislukukausi ja -vuosi: Syksy 2014 Sivumäärä: liitettä CRISPR on uusi tarkka menetelmä geenitekniikan alalla. Se perustuu alun perin bakteerien virustorjuntaan, jossa viruksen DNA:ta lisätään osaksi bakteerin omaa genomia. CRISPR on luotettava, melko edullinen ja yksinkertainen geeninmuokkausmenetelmä. Työn tarkoituksena oli luoda poistogeeninen solulinja CRISPR-menetelmän avulla. Geeni, joka työssä pyrittiin poistamaan, vastaa AE2-proteiinin tuotannosta Golgin laitteessa. AE2-proteiinin tehtävänä solussa on siirtää ainioneita solukalvon läpi ja säädellä Golgin laitteen ph:ta. Proteiini on todennäköisesti soluille tärkeä ph:n ylläpidossa, ja solulinjan luomisella haluttiin todistaa sen merkitys niille. Solulinjan valmistamisessa ei onnistuttu, vaikka klooneja onnistuttiinkin saamaan työssä 4 kpl. On mahdollista, että proteiini AE2, jonka tuotannon poistogeenisyys lopettaa, oli soluille liian tärkeä elämisen kannalta ja onnistuneen transfektion solu kuolivat. Toisena syynä tähän saattoi olla plasmidin ihmisperäisyyden ja solulinjan apinaperäisyyden yhteensopimattomuus. Tästä johtuen sekvenssissä kriittisessä kohtaa sijainnut yhden emäsparin eroavaisuus saattoi estää halutun poistogeenisen DNA-palan siirtymisen solun genomiin. Työ tehtiin Oulun yliopiston biokemian ja molekyylilääketieteen tiedekunnassa syksyllä 2014 Sakari Kellokummun ryhmässä. Tämä oli ensimmäinen kerta, kun tätä geeninmuokkausmenetelmää käytettiin kyseisessä ryhmässä. Asiasanat: CRISPR, geenitekniikka, kloonaus, soluviljely 3
4 SISÄLLYS TIIVISTELMÄ 3 SISÄLLYS 4 1 JOHDANTO 4 2 ELÄINSOLU Tuma ja nukleiinihapot DNA RNA Golgin laite Tehtävä AE2-proteiini 8 3 CRISPR JA MUUT KÄYTETYT MENETELMÄT CRISPR Alkuperä CRISPR menetelmän käyttö eläinsolussa PCR ja agaroosigeelielektroforeesi Sekvensointi Transfektio Virtaussytometri Immunofluoresenssivärjäys 15 4 ELÄINSOLUJEN VILJELY Aseptinen työskentely Eläinsoluviljely Eläinsolujen jakaminen uudelle alustalle Solulinjat 19 5 TYÖN KOKEELLINEN OSUUS Plasmidin lisäys COS 7 -solut Genomisen DNA:n eristys ja PCR Sekvensointi 23 4
5 5.5 Transfektio Sorteeraus 24 6 KLOONIEN KASVATUS JA TARKISTAMINEN Kloonien kasvatus Immonofluoresenssivärjäys Mikroskopia 27 7 TULOKSET JA POHDINTA 28 8 YHTEENVETO 30 LÄHTEET 32 5
6 1 JOHDANTO Eläinsolut kuuluvat eukaryoottisiin eli aitotumallisiin soluihin. Ne ovat hyvin monipuolinen ja erilaistunut solutyyppi ja poikkeavat näin ollen muista eukaryootteista eli hiiva- ja kasvisoluista. Eläinsolut koostuvat useista eri soluelimistä. Näitä ovat esimerkiksi solukalvo, mitokondrio, tuma ja Golgin laite. Työn tavoitteena oli luoda poistogeeninen solulinja CRISPR-menetelmää käyttäen. CRISPR-menetelmä on vuonna 2013 julkaistu geeninmuokkausmenetelmä, joka perustuu bakteerien puolustusmekanismiin viruksia vastaan. Menetelmä mahdollistaa DNA:n tarkan muokkaamisen, ja se on huomattavasti luotettavampi kuin edelliset muokkausmenetelmät. CRISPR-menetelmällä pystytään muokkaamaan elävien solujen genomia ja sillä onkin saatu lupaavia tuloksia laboratoriokokeissa geneettisten sairauksien osalta. Geeni, jonka poistaminen työn tavoitteena oli, vastaa Golgin laitteessa sijaitsevasta AE2-proteiinin tuotannosta. Tämän proteiinin tehtävänä on solukalvolla ja Golgin laitteessa vastata kloridi- ja bikarbonaattianioneiden kuljettamisesta solukalvon läpi ja siten solun ja Golgin laitteen ph:n säätelystä. Geenin poistamisella solusta haluttiin nähdä, mitä vaikutuksia kyseisen proteiinin puuttuminen aiheuttaa Golgin laitteen toiminnassa, erityisesti sen sokerirakenteiden muokkauksessa. Työ tehtiin Oulun yliopiston Biokemian ja molekyylilääketieteen tiedekunnassa syksyllä 2014, ja sen tavoitteena oli saada aikaiseksi poistogeeninen solulinja jatkotutkimuksia varten. 4
7 2 ELÄINSOLU Eläinsolut kuuluva eukaryootteihin eli aitotumallisiin soluihin, joita ovat hiivat, homeet ja kasvisolut. Niillä on useita soluelimiä, kuten solukalvo, solulima, mitokondrio, ribosomit, solulimakalvosto, tuma ja Golgin laite. (Kuva 1.) Eläinsoluja ympäröivä ohut solukalvo muodostuu kaksoiskerroksisesta lipidikalvosta. Siihen on myös kiinnittynyt kolesterolia ja useita eri proteiineja, joista monissa on liittyneenä sokeriketjuja. Ohuen solukalvon takia eläinsolut ovat varsin hauraita ja ne vaurioituvat herkästi mekaanisen rasituksen voimasta. (1, s ; 2; 3.) Solulima on solun välitysaine, jossa osa biokemiallisista reaktioista tapahtuu. Se on suurimmaksi osaksi vettä, mutta siinä on myös lipidejä, epäorgaanisia ioneita ja proteiineja. Proteiinien synteesi soluissa tapahtuu ribosomien pinnalla. Nämä pienet soluelimet koostuvat RNA:sta ja proteiineista. Ne kelluvat yleensä solulimassa, mutta saattavat olla myös kiinnittyneenä solulimakalvostoon. Solulimakalvostossa valmistetaan ja muokataan proteiineja ja lipidejä. Se myös osallistuu aineiden, kuten mitokondrion muokkaamien orgaanisten molekyylien kuljetukseen. Mitokondrio vastaa soluissa energia-aineenvaihdunnasta. Sille kuuluu niin kemiallisen energian muuttaminen käyttökelpoiseen muotoon kuin soluhengityskin. (2; 4, s. 70, 78.) KUVA 1. Eläinsolun rakenne (5) 5

8 2.1 Tuma ja nukleiinihapot Tuma on eukaryoottisoluissa perintöaineksen sisältävä soluorganelli. Se rakentuu kaksinkertaisen kalvon muodostamasta tumakotelosta, tumahuokosista, jotka mahdollistavat nukleiinihappojen ja proteiinin liikkumisen solussa sekä tumaplasmasta, jossa solun perintöaines eli geenit sisältävä kromatiini lepäävät. Kromatiini koostuu pakkautuneesta kaksoiskierteisestä DNA-rihmastosta, joka on kiertynyt histoni-proteiinien ympärille. (4, s. 74; 6.) DNA DNA on kaksijuosteinen kierteinen nukleiinihappo, joka koostuu lukuisista toisiinsa liittyneistä nukleotideista. Nukleotidit muodostuvat neljästä erilaisesta emäksestä (adeniini, tymiini, guaniini ja sytosiini) sekä sokeri- ja fosfaattiosasta (kuva 2). (7; 8, s ) KUVA 2. RNA:n ja DNA:n rakenne (9) 6
. RNA:ta tuotetaan DNA:sta ja RNA välittää informaation proteiinien tuottoa varten. (8, s. 15 22.) 2.")
9 2.1.2 RNA RNA koostuu DNA:n tavoin nukleotideistä, mutta sen rakenne on yksijuosteinen ja se on yleensä kooltaan pienempi molekyyli kuin DNA. RNA:n poikkeaa myös emäskoostumukseltaan DNA:sta. Siinä tymiini-emäksen tilalla on urasiili-niminen emäs (kuva 2). RNA:ta tuotetaan DNA:sta ja RNA välittää informaation proteiinien tuottoa varten. (8, s ) 2.2 Golgin laite Golgin laite on soluelin, joka sijaitsee eläinsoluissa yleensä tuman lähellä. Se löydettiin vuonna 1898, jonka jälkeen tutkijat ovat pyrkineet selvittämään tämän soluelimen tarkoitusta ja rakennetta. Kuitenkaan Golgin laitteen tapaa kuljettaa muokattavia proteiineja ei vieläkään tunneta täysin. (10, s. 21) Golgin laite rakentuu 5 10:stä kaksikerroksen lipidikalvon ympäröimästä ontelosta eli kisternoista (kuva 3). Kisternat ovat asettuneet päällekkäin ja muodostavat matalaa kupolia muistuttavan pussipinkan. Golgin laite erotellaan kahteen puoleen, cis- ja trans-puoleen. Cis-puoli sijaitsee kupolin kuperalla ja trans-puoli kupolin koveralla puolella. Molekyylit liikkuvat Golgin laitteessa cispuolelta trans-puolelle. (10, s. 21; 11; 12, s.75.) KUVA 3. Golgin laitteen rakenne (13) 7
10 2.2.1 Tehtävä Golgin laitteen tehtävänä on muokata proteiineja ja lipideja lisäämällä niihin hiilihydraattiosia eli sokereita. Proteiinien liikkumiseen Golgin laitteessa on kaksi eri teoriaa. Ensimmäisen mukaan proteiinit liikkuvat kisternoista toiseen vesikkeleissä eli kalvostosta irtoavissa rakkuloissa. Toisen teorian mukaan kisternat kulkeutuvat hitaasti eteenpäin cis-puolelta trans-puolelle ja kypsyvät siinä samalla. Päästyään pinon päällimmäiseksi kisterna jakautuu useiksi vesikkeleiksi ja vesikkelit kulkeutuvat muualle soluun. Samalla pinon alimmaksi muodostuu aina uusi kisterna. Ei tiedetä, kumpi teorioista on oikea, ja on mahdollista, että Golgin laitteistossa käytetään molempia tapoja: nopeaa vesikkeleissä kulkemista ja hidasta kirsternojen kulkeutumista. (10, s. 24; 12, s 75.) Eri kisternoissa vallitsevat erilaiset olosuhteet aina ph:sta entsyymeihin. Näin jokaisessa kisternassa tapahtuva proteiinin muokkaus on erilaista kuin toisessa. Valmiit proteiinit Golgin laite erittää ympäröivään solulimaan kalvorakkuloissa, joista ne kulkeutuvat ympäri solua. Muina tehtävinä Golgin laitteella on lajitella proteiineja solukalvolle, lysosomeihin tai solun ulkopuolelle sekä valmistaa kalvomateriaalia solun käyttöön. (10, s. 24; 11; 12, s. 333.) AE2-proteiini Proteiinit ovat elintoiminnoille välttämättömiä valkuaisaineita, joilla on useita eri tehtäviä soluissa. Ne voivat toimia entsyymeinä, jolloin ne toimivat solun sisällä katalysaattoreina useille kemiallisille reaktioille. Proteiinit voivat myös sitoa solulle tärkeitä aineita tai osallistua aineiden kuljetukseen. Ne myös toimivat viestinviejinä eli hormoneina toisille soluille, esimerkiksi kasvuhormonina. Osa proteiineista toimii myös solujen tukirakenteina, kuten tubuliini tai aktiini. (14.) Primaarirakenteeltaan proteiinit koostuvat peptidisidoksin toisiinsa kiinnittyneistä aminohapoista eli polypeptidiketjuista. Nämä ketjut poimuttuvat tai kiertyvät serpentiinimäiseksi kierteeksi muodostaen vetysidoksien kanssa sekundaarirakenteen. Kun primaari- ja sekundaarirakenteet sekoittuvat 8
11 keskenään ja niiden aminohapot muodostavat toisiinsa erilaisia sidoksia, puhutaan kolmiulotteisesta tertiaarirakenteesta. Kvartaarirakenne eli lopullinen proteiinimolekyyli muodostuu tällöin useista tertiäärirakenteisista aminohappoketjuista. (14; 15.) AE2-proteiini (Anion Exchanger) sijaitsee Golgin laitteessa. Sen tehtäviä ovat toimiminen anioneiden kuljetusproteiinina solukalvolla, säädellä Golgin laitteen ph:ta ja lisätä veren kykyä kuljettaa hiilidioksidia keuhkoihin. AE2-proteiini on todennäköisesti soluille merkittävä proteiini, koska sen puuttuessa soluihin kertyy liikaa bikarbonaattianioneita. Tämä puolestaan aiheuttaa solun ph:n tason nousun ja vaikuttaa siten solun kykyyn lisätä sokerirakenteita proteiineihin ja lipideihin. Pahimmillaan ph:n nousu voi aiheuttaa jopa solukuoleman. (16; 17.) 9
12 3 CRISPR JA MUUT KÄYTETYT MENETELMÄT Geenitekniikka on biotekniikan osa, joka keskittyy geneettiseen muunteluun. DNA:n samankaltainen rakenne jokaisella eliölajilla ja nukleotidijärjestyksen tietäminen on mahdollistanut suunnitellun geneettisen muuntelun. Nykyisin käytettyjä menetelmiä geenin muokkaukseen ovat TALENs (Transcription activatorlike effector nucleases), sinkkisormiproteiinit ja CRISPR. TALENs menetelmässä genomista poistetaan pala. Sinkkisormiproteiinimenetelmä on samankaltainen kuin TALENs, mutta siinä sinkkisormet sitoutuvat katkaistavaan kohtaan ja katkaisevat genomin. Molemmat menetelmät ovat laajasti käytettyjä, mutta eivät yhtä tarkkoja ja luotettavia menetelmiä kuin uusi CRISPR-menetelmä. (18; 19.) 3.1 CRISPR CRISPR (clustered regularly interspaced short palindromic repeats) on geenimuokkaustekniikka, jolla voidaan muokata elävien solujen genomia. Se perustuu bakteerien virusinfektioiden puolustusjärjestelmään ja sen kopioimiseen muissa soluissa. (20.) Alkuperä Bakteerien puolustusmekanismi viruksia vastaan perustuu siihen, että bakteerit lisäävät itseensä hyökänneen viruksen DNA:sta osia omaan genomiinsa. Ne kääntävät vieraan DNA:n RNA:ksi, pilkkovat RNA:n palasiin sekä liittävät paloihin guide- eli ohjain-rna:n. ja CRISPR-entsyymit. Tämän jälkeen ne muodostavat yhdessä Cas9-entsyymin kanssa kompleksin. Cas9-entsyymi etsii ohjain- RNA:n mukaisen kohdan bakteerin DNA:sta ja irrottaa juosteet toisistaan. Sen jälkeen CRISPR-entsyymit leikkaavat yksijuosteisesta DNA:sta ohjain-rna:n määrittelemän palan pois. Tässä vaiheessa bakteerin korjaajaentsyymit alkavat nopeasti korjata rikkoutunutta DNA:ta ja kääntävät kuljetetusta vieraasta RNA:sta ketjuun uuden DNA-sekvenssin. (20; 21.) 10
13 3.1.2 CRISPR menetelmän käyttö eläinsolussa CRISPR-menetelmässä solujen sisälle viedään plasmidi, johon on liitetty haluttu maksimissaan 20 emäsparin pituinen sekvenssi, ohjain-rna sekä CRISPR-entsyymien RNA:t, jotka vastaavat Cas9-enstyymien ja reportteriproteiinin (fluoresoiva EGFP) muodostuksesta (kuva 4). KUVA 4. AE2 Clone 1 P x 458-plasmidin rakenne (22) Solu alkaa tuottaa plasmidista Cas9-entsyymiä ja EGFP-proteiinia RNA:n ohjeen mukaan. Syntyneet Cas9-entsyymit muodostavat plasmidista kopioidun ohjain-rna:n kanssa kompleksin, joka etsiytyy oikeaan kohtaan solun genomissa (kuva 5). EFGP-proteiini jää loistamaan hetkeksi solun sisään merkkinä siitä, että solu on ottanut halutun plasmidin sisäänsä. Myöhemmin solu poistaa 11
14 itsestään tämän plasmidin, jolloin sekä Cas9-entsyymin että EFGP-proteiinin tuotto loppuu. (21; 23.) KUVA 5. CRISPR-menetelmä eläinsolussa (24) 3.2 PCR ja agaroosigeelielektroforeesi PCR eli polymeraasiketjureaktio (eng. polymerase chain reaction) on menetelmä, jolla pystytään joustavasti monistamaan DNA-jaksoja. Se perustuu monistettavan alueen rajaaviin alukkeisiin ja korkeita lämpötiloja sietäviin DNA-polymeraaseihin, jotka löydettiin kuumissa lähteissä elävistä mikrobeista. Menetelmän keksi Kary Mullis vuonna 1983, ja se on nykyisin yleisesti käytössä eri alojen laboratorioissa. PCR-työskentely vaati erityistä huolellisuutta pienten pipetointimäärien vuoksi. Lisäksi siinä on huomioitava tarkkaan oikeat lämpötilat, ohjelman aikataulutus sekä lähtö-dna:n puhtaus. Esimerkiksi veri sisältää kopiointia häiritseviä tekijöitä. PCR:ssä pitää myös varoa näytteen hajoamista. (8, s ; 25.) 12
15 PCR-tekniikka perustuu kolmeen vaiheeseen. Ensimmäisessä vaiheessa eli denaturaatiossa reaktioseoksen lämpötila nostetaan korkealle (esim. 95 o C:seen), jolloin DNA denaturoituu eli juosteet irtoavat toisistaan. Toisessa vaiheessa lämpötilaa lasketaan (usein o C:seen) hetkellisesti niin, että alukkeet voivat sitoutua yksijuosteisiin DNA-molekyyleihin. Tätä vaihetta kutsutaan annealing-reaktioksi. Viimeisessä vaiheessa lämpötilaa jälleen nostetaan (esim. 72 o C:seen) DNA-polymeraasin toiminnalle suotuisaksi, jolloin DNA-syntetisoi yksijuosteisesta DNA:sta kaksijuosteista DNA:ta. Viimeistä vaihetta kutsutaan elongaatio-reaktioksi. Näitä kolmea vaihetta eli sykliä toistetaan, kunnes saadaan monistettua haluttu määrä DNA:ta. Viimeisen syklin elongaatio on yleensä pidempi, jotta kaikki DNA:n yksijuosteet saataisiin varmasti syntetisoitua kaksijuosteiseksi. (8, s ; 25.) 3.3 Sekvensointi Sekvensoinnilla tarkoitetaan DNA:n nukleiinihappojärjestyksen selvittämistä. Menetelmiä käytetään yleisesti esimerkiksi mutaatioiden havainnoinnissa tai sukulaisuuden selvittämisessä. Sekvensointiin on käytössä kaksi eri menetelmää, Sangerin menetelmä ja Maxam-Gilbertin menetelmä. Nykyisin sekvensoinnit suoritetaan lähes aina Sangerin menetelmällä sen myrkyttömyyden vuoksi. Menetelmä perustuu DNA-juosteiden aukeamiseen kuumennuksen johdosta, jolloin niihin liitettään jokin neljästä diseoksinukleotidista. Tämän jälkeen DNA jaetaan 4 eri koeputkeen, joista jokaisessa on vain yhtä tiettyä diseoksinukleotidia. Diseoksinukleotidin tehtävänä on pysäyttää DNA:n kahdentumisreaktio ja pilkkoa DNA paloihin aina tietyn emäksen kohdalta, esimerkiksi tymiinin. Kun näytteet tämän jälkeen erotellaan elektroforeesilla, saadaan selville DNA:n sekvensin emäsjärjestys. Myös sekvensointilaitteet perustuvat useimmiten Sangerin menetelmään. (26; 27.) 13
16 3.4 Transfektio Transfektiolla tarkoitetaan DNA:n siirtymistä joko luonnollisesti tai keinotekoisesti elävän eläinsolun sisälle. Transfektion tulos voi olla joko pysyvä tai väliaikainen ekspressio eli geenin ilmentyminen. Pysyvässä eli stabiilissa ekspressiossa DNA integroituu isäntäsolun genomiin ja siirtyy solun jakautuessa tytärsoluun. Väliaikaisessa eli transientissa ekspressiossa ominaisuus katoaa solun jakautuessa. (8. s ; 28, s.17.) Eläinsolujen transfektioon on kehitetty lukuisia menetelmiä, ja monet näistä menetelmistä ovat spesifisiä tietyille solulinjoille. Yksi laboratoriossa suosittu menetelmä on liposomimenetelmä. Siinä DNA viedään solun sisälle pienten lipidirakkuloiden sisällä, koska DNA on hydrofiilinen ja ei sen takia pääse hydrofobisen solun sisään itsestään. Positiivisesti varautuneet kationilipidit ympäröivät DNAmolekyylin, joka on fosfaattien takia on negatiivisesti varautunut. DNA-lipidikompleksin osuessa soluun lipidikerros joko sulautuu solukalvoon ja työntää DNA:n solun sisälle tai menee solukalvon läpi endosytoosin avulla. Muita ylesesti käytettyjä menetelmiä ovat mm. kalsiumfosfaattikopresipitaatio ja mikroinjektio. Viimeksi mainittua voidaan käyttää ainoastaan hyvin suurikokoisille soluille esim. hedelmöittyneille munasoluille. (8, s ; 28, s.17.) 3.5 Virtaussytometri Solujen sortteeraus eli lajittelu perustuu virtaussytometriaan. Menetelmässä solut pakotetaan kulkeutumaan yksitellen virtauskammioon ja lasersäteen ohitse. Tietyn aallonpituuden omaavan lasersäteen osuessa fluoresoivilla leimoilla merkattuihin soluihin (yleensä käytetään aallonpituutta 488 nm) fluorokromi virittyy ja melkein välittömästi purkautuu eli alkaa lähettää virittäneen laserin aallonpituutta pidempää valoa. Tätä purkautuvaa valoa ohjataan sitten halutulle detektorille peilien ja suodattimien läpi. Detektoriin tulevan valon avulla solut voidaan jakaa spesifisimpiin ryhmiin. Laitteen sisällä pieni anturi lajittelee halutut solut 14
17 keräysputkeen. Sortteerauksessa käytettävällä virtaussytometrilla on lisäominaisuus, jolla pystytään jakamaan solut yksitellen 96-kuoppalevyn kuoppaan. ( 28, s. 17; 29.) 3.6 Immunofluoresenssivärjäys Immunofluoresenssivärjäyksellä tarkoituksena on osoittaa fiksatuista soluista tai kudoksista haluttuja proteiineja tai antigeenejä käyttämällä fluoresoivalla merkkiaineella eli fluorokromilla merkittyjä vasta-aineita. Fluorokromeina käytetään fluoresenssimikroskopiassa yleisesti mm. fluoreseiiniä ja rodamiiniä, koska ne absorboivat lyhytaaltoista valoa. Samassa värjäyksessä voidaan osoittaa useita eri antigeenejä tai proteiineja käyttämällä eri väreissä fluoresoivia sekundaarivasta-aineita. (29.) Immunofluoresenssivärjäyksestä on kaksi menetelmää, suora ja epäsuora. Molemmissa menetelmissä värjättävä näyte blokataan ensin proteiinilla, esimerkiksi BSA:lla (Bovine serum albumin eli albumiini-proteiinia naudan seerumista), jotta vasta-aineet sitoutuvat vain oikeisiin kohteisiin. Blokkaaminen myös vähentää taustan värjäytymistä, jolloin proteiinit tai antigeenit erottuvat paremmin materiaalista. Suorassa menetelmässä käytetään ainoastaan yhtä vasta-ainetta. Se sitoutuu suoraan tarkoitettuihin antigeeneihin tai proteiineihin. (30; 31) Epäsuorassa menetelmässä käytetään kahta erilaista vasta-ainetta. Ensimmäiseksi materiaalille laitetaan primäärivasta-aine, jonka tehtävänä on sitoutua antigeeniin tai proteiiniin kuten suorassa menetelmässäkin. Sen jälkeen materiaalille laitetaan fluoresoiva sekundaarivasta-aine, jonka tehtävä on taas sitoutua primaarivasta-aineeseen. Molemmat menetelmät kuitenkin näyttävät samalla tavalla materiaalista värjäystuloksen. Epäsuora menetelmä on herkempi kuin suora menetelmä, koska siinä useampi fluorofori sitoutuu samaan paikkaan. (Kuva 6.) (30; 31.) 15
18 KUVA 6. Suora ja epäsuora immunofluoresenssivärjäys (31) 16
19 4 ELÄINSOLUJEN VILJELY Soluviljelyn avulla soluja voidaan tutkia tarkasti kontrolloiduissa olosuhteissa. Laboratorio-olosuhteissa kasvaakseen solut tarvitsevat tarkkaan säädellyt ympäristön ravintoaineet, lämpötilan ja ph-arvo. Lisäksi soluviljely vaatii aseptista työskentelyä. (32.) 4.1 Aseptinen työskentely Soluviljelyssä noudatetaan aseptista työskentelyä, jotta mahdolliset soluille haitalliset bakteeri-, sieni- ja mykoplasmakontaminaatiot voitaisiin estää tehokkaasti. Aseptisen työskentelyn periaatteena on myös suojata solulinjoja ristikontaminaatioilta sekä työntekijöitä työskentelyyn liittyviltä riskeiltä. Soluviljely tapahtuu puhdastilassa eli suljetussa tilassa, johon tuleva ilma suodatetaan HEPA-suodattimilla sekä tilan ylipaineistuksella. Puhdastilassa työntekijä pukee ylleen suojavaatetuksen, johon kuuluvat laboratoriotakki, työjalkineet, kertakäyttöiset hanskat ja mahdolliset kasvo- ja hiussuojukset. (33.) Työskentely tapahtuu laminaarivirtauskaapissa, joka voi olla suljettu tai avoin pysty- tai vaakavirtauskaappi. Siinä ilmavirtaus estää huoneilman pääsemisen kaapin sisälle, jolloin kaapin sisäinen työtila pysyy steriilinä ja suojaa työntekijää käsiteltäviltä materiaaleilta. Laminaarin sisälle vietävät esineet tulee desinfioida hyvin 70-prosenttisella alkoholilla, jotta laminaarin sisäpuoli pysyisi steriilinä. (33.) 4.2 Eläinsoluviljely Eläinsolut kasvavat laboratoriossa normaalisti kiinteillä pinnoilla, koska kudoksissa olevat solut ovat tiiviisti kontaktissa toisiinsa erilaisten soluliitosten välityksellä. Alustoina eläinsoluja viljeltäessä käytetään yleensä lasipintoja tai käsiteltyä muovia, jonka pinnalla on negatiivisesti varautuneita ryhmiä. Solut erittävät viljelyastian pintaan kollageeniä ja muita soluväliaineen komponentteja, joilla ne 17
20 sitoutuvat kiinni viljelyastian pintaan. Näille alustoille solut muodostavat yksisolukerroksen ja alkavat kuolla kerroksen täytettyä kasvualustan kokonaan. (32; 34.) Alustojen pinnalla käytetään tarkkaan määriteltyä kasvatusliuosta, josta solut saavat ravinteensa. Soluille tarkoitettu kasvatusliuos sisältää runsaasti epäorgaanisia suoloja, aminohappoja, vitamiineja, glukoosia ja typpilähteen. Epäorgaanisilla suoloilla, kuten kalium, magnesium, natrium, kloridi ja fosfaatti, säädetään liuoksen happamuus ja osmoottinen paine soluille sopivaksi. Aminohapoista liuos sisältää eniten glutamiinia, koska nisäkässolut käyttävät sitä glukoosin lisäksi hiilen lähteenä ja energiana. Kasvatusliemeen lisätään myös usein seerumia 5 20 prosenttia tilavuudesta. Seerumi sisältää aminohappoja, hivenaineita, proteiineja, lipidejä, vitamiineja, hormoneja ja kasvutekijöitä. Seerumin käyttö lisää solujen jakautumista, sitoo raskasmetalleita ja suojaa nisäkässoluja. Kasvatusliuos sisältää myös eri antibiootteja, kuten penisilliiniä bakteereita ja streptomysiiniä hiivoja vastaan. (32; 34.) 4.3 Eläinsolujen jakaminen uudelle alustalle Eläinsolut jaetaan uudelle alustalle normaalisti siinä vaiheessa, kun sen hetkinen alusta alkaa olla kasvanut täyteen. Alustaa ei saa kuitenkaan päästää kasvamaan aivan täyteen, etteivät solut ala kuolemaan tilan tai ravinteiden loputtua. Myös kasvatusliemen ph:n lasku aiheuttaa soluissa muutoksia ja pahimmillaan kuoleman. Aluksi jaettavista soluista poistetaan kasvatusliemi ja ne pestään sopivalla pesuliuoksella (PBS lähes aina). Tämän jälkeen solujen päälle tiputetaan juuri kasvatusalustan pohjan peittävä määrä joko trypsiiniä tai EDTA:ta, joiden tarkoituksena on irrottaa solut kasvatusalustasta. Irtoamista varten kasvatusalustat laitetaan 1 2 minuutiksi + 37 o C:seen. Irrotettaessa ajan kanssa kannattaa olla varovainen, jottei trypsiini tai EDTA pääse hajottamaan myös solukalvoa, jolloin solu kuolee. (34.) 18
21 Irrottamisen jälkeen kasvatusalustalle lisätään kasvatuslientä, joka neutralisoi trypsiinin tai EDTA:n hajottavan vaikutuksen. Solut sisältävä liuos voidaan siirtää falcon-putkeen, jossa solut sentrifugoidaan varovasti putken pohjalle. Kierrosnopeus pidetään matalana, jotteivät solut hajoaisi rasituksen vaikutuksesta. Kun solut on saatu painumaan pohjaan, poistetaan liuos niiden päältä ja solut suspensoidaan kasvatusliemeen. Eläinsolut voidaan myös siirtää suoraan irrotuslioksessa uudelle kasvualustalle. Tämä ei kuitenkaan ole suotavaa irrotusliuoksen soluja hajottavan ominaisuuden vuoksi. Pieninä määrinäkin se voi heikentää solujen jakautumista. Yleensä solut lasketaan suspensoidusta liuoksesta, jotta tiedettäisiin, kuinka paljon niitä siirretään uudelle kasvatusalustalle kasvamaan. Lopuksi lisätään solujen päälle kasvatusliemi ja kasvatusalustat siirretään + 37 o C:seen CO2-inkubaattoriin kasvamaan. (34.) 4.4 Solulinjat Solulinjalla tarkoitetaan jatkuvasti jakautuvaa solupopulaatiota, jonka elinikä on rajaton. Pysyvät eläinsolulinjat ovat usein eristetty kasvainkudoksista tai geneettisesti muunnellusta primaarisoluviljelmästä, joka on transformoitunut kemiallisten mutageenien, säteilyn tai virusten vaikutuksesta. Sen vuoksi ne ovat alkaneet käyttäytyä kasvainsolumaisesti. (35; 36.) Tällaiset nopeakasvuiset ja kuolemattomat eläinsolulinjat sopivat hyvin tutkimusmateriaaliksi. Niillä on kuitenkin geneettisestä muuntelusta johtuvia poikkeavia ominaisuuksia, kuten epänormaali kromosomilukumäärä. Myös niiden kasvukontrolli poikkeaa normaalista, ja ne voivat kasvaa ilman alustaan kiinnittymistä. Tämän vuoksi ne myös voivat aiheuttaa kasvaimia päästessään isäntäorganismiinsa ja aiheuttavat sen vuoksi turvallisuusriskin. (35; 36) Tunnetuin varsin erilaistumaton solulinja on amerikkalaisen Henrietta Lacksin kohdunkaulansyövästä eristetty HeLa-solulinja. HeLa-soluja käytetään laboratorioissa paljon perustutkimukseen esimerkiksi solun rakenteiden, DNA:n tai RNA:n tutkimiseen. (35; 36.) 19
22 5 TYÖN KOKEELLINEN OSUUS Työ ensimmäisessä vaiheessa valittiin muokattava solulinja ja tuotettiin lisää plasmidia. Seuraavassa vaiheessa plasmidi tarkistettiin solulinjaan sopivaksi ja transfektoitiin soluihin. Viimeisessä vaiheessa transfektoidut solut lajiteltiin kasvamaan yksittäin ja eloon jääneet tarkistettiin. Työ suoritettiin kesällä ja syksyllä 2014 Oulun yliopiston Biokemian ja molekyylilääketieteen tiedekunnassa. 5.1 Plasmidin lisäys Työn aluksi lisättiin plasmidin AE2 Clone 1 P x 458 määrää. Lisäämistä varten plasmidi oli siirrettävä bakteerin sisälle, josta se voitiin eristää riittävän bakteerikasvun jälkeen. Plasmidi transformoitiin kaupallisiin XL1 Blue -kompetenttisoluihin ohjeen mukaan (liite 2). DNA:n pitoisuus transformaatiossa oli 570 ng/µl. Transformoidut solut laitettiin kasvamaan LB-ampisilliinimaljalle yön yli. Seuraavana päivänä siirrostettiin silmukalla maljalle kasvaneista pesäkkeistä yksittäinen pesäke uudelle LB-ampisilliinimaljalle ja lopuksi silmukkaa heiluteltiin 4 ml:ssä LB-ampisilliinilientä. Näin saatiin siirrettyä saman pesäkkeen bakteereja myös testikasvustoon. Edellisen päivän 4 ml:n kasvatuksesta tehtiin mini-prep eli eristettiin plasmidi E.Z.N.A Plasmid DNA Mini Kit I -kitillä. Eristyksellä tarkistettiin, että transformoitu DNA on säilynyt entisellään ja sen kloonaaminen on onnistunut. Tämän jälkeen DNA:sta tehtiin digestio ohjeen mukaan (liite 2) ja digestioista laitettiin agaroosigeelille ajo tulemaan. Ajossa käytettiin markkerina kaupallista Thermo Scientificin GeneRuler 1 kb DNA Ladderia. Geeli ajettiin 300 V:n teholla, jolloin se kesti vain 22 minuuttia. Ajosta näkyi, että kloonattu DNA oli onnistuttu säilyttämään. Sen johdosta laitettiin DNA:n suurempaa eristystä varten 1-1-kloonia kasvamaan 100 ml:aan LB-lientä, johon oli lisätty antibioottina 1ml ampisilliinia. Midi-prep eli DNA:n suurempi eristys tehtiin Nucleobond Xtra Midi -kitillä. Sen avulla saatiin eristettyä melko puhdasta DNA:ta 1,8 µg/µl, mikä on hyvä saanto. Puhdistetut ja käyttövalmiit kloonit pakastettiin odottamaan tulevaa transfektiota. 20
23 5.2 COS 7 -solut Työssä käytettiin kaupallista COS 7 -solulinjaa, joka on eristetty apinan munuaisesta ja transfektoitu SV40-viruksella. Transfektion takia solut voivat jakautuvat loputtomasti. Yleensä niiden käyttö kuitenkin lopetetaan jakamiskerran jälkeen, koska silloin solulinja on ehtinyt muuttua liikaa alkuperäisistä soluista. COS 7 -solut ovat jakautumisnopeudeltaan melko hyviä. Valintaan vaikutti myös solujen aiempi käyttö kyseisessä tutkimuksessa ja näin ollen tulevien tulosten mahdollinen liittäminen vanhoihin tutkimustuloksiin. Työhön mietittiin myös käytettäväksi ihmisen ihon fibroblasti-primaarisolulinjaa, mutta se hylättiin solujen hitaan jakautumisnopeuden takia. 5.3 Genomisen DNA:n eristys ja PCR COS 7 -solujen genomisen DNA:n selvittämiseksi piti soluista ensiksi eristää DNA ja monistaa sitä lisää nukleotidijärjestyksen selvittämistä varten. Järjestys oli tärkeä saada selville, jotta nähtäisiin, missä kohdin genomia transfektoitava plasmidivektori vaikuttaisi. Aluksi COS 7 -soluja jaettiin kahdelle 10 cm:n maljalle kasvamaan. Kahden päivän kuluttua irrotettiin hyvin jakautuneet solut 2 ml:lla 2 x trypsiiniä, jonka annettiin irrottaa soluja 10 minuutin ajan. Soluihin lisättiin trypsiinin neutraloimiseksi DMEM-kasvatuslientä, siirrettiin solut ja liuokset 15 ml:n falcon-putkeen ja sentrifugoitiin solut pohjaan 4 minuuttia 1000 x g voimalla. Solupelletin päältä poistettiin erottunut neste ja solupelletti suspensoitiin 900 µl:aan DMEM-lientä. Suspenssioista otettiin 5 µl 1,5 ml:n eppendorf-putkiin ja lisättiin joukkoon 50 µl DNA Extraction Solution -liosta. Putkia sekoitettiin 15 sekuntia Vortex-laitteella. Tämän jälkeen putket laitettiin inkuboitumaan + 65 o C:n lämpöblokkiin 15 minuutiksi. Lämpöblokin lämpötilaa nostettiin + 68 o C:seen ja annettiin solujen vielä inkuboitua 15 minuutin ajan. Putket siirrettiin toiseen + 98 o C:n lämpöblokkiin ja annettiin olla siinä 10 minuuttia. Kaiken jälkeen sakka sentrifugoitiin putken pohjaan 700 rpm:n voimalla pöytäfuugissa minuutin ajan. 21
24 PCR-reaktioita varten reaktioliuokset pipetoitiin pieniin eppendorf-putkiin taulukon 1 mukaan. TAULUKKO 1. Pipetointi ohje PCR-reaktiota varten PCR - reaktio yht. 50 µl X 4 kpl Liuos Määrä ( µl ) Muuta Eristetty genominen DNA - liuos 3 Forward - primer 10 µm 1,25 Reverse - primer 10 µm 1,25 dntp - mix 1 Fermetas - merkkinen PCR - puskuri 5 sisältää MgCl iproof - PCR entsyymi 0,5 Pidettävä kylmänä ja lisätään viimeisenä ennen vettä. de. ion. H 2 O 38 Pipetoinnin jälkeen putket käytettiin minuutin ajan sentrifugissa, jotta kaikki liuokset saatiin painumaan pohjalle ja sekoittumaan keskenään. Tämän jälkeen putket asetettiin PCR-laitteeseen. Laitteelle syötettiin taulukon 2 mukainen ohjelma. TAULUKKO 2. PCR-laitteelle syötetty ajo-ohjelma PCR - ohjelma Vaihe Lämpötila ( o C ) Aika Denaturaatio 98 2 min Sykli Denaturaatio s x Annealing s 35 krt Elongaatio s Loppu 72 3 min Odottamistila 4 - PCR-laitteen toimiessa tehtiin DNA:n tarkistusta varten 1,5 prosettinen agaroosigeeli. Geeliä tehtiin 100 ml:n annos, johon tuli 1,5 g agaroosijauhetta ja 5 µl etidiumbromideja. Aluksi agaroosijauhe keitettiin mikrossa deionisoidussa vedessä, minkä jälkeen siihen lisättiin sekaan etidiumbromidia. Agaroosigeelin annettiin jäähtyä kädenlämpöiseksi ja kaadettiin se valukelkalle. Valukelkalla geelistä puhkottiin neulalla kaatamisessa syntyneet ilmakuplat ja siihen asetettiin 10 kaivon kampa. Annettiin geelin jähmettyä ja poistettiin kaivokampa varovasti. 22
 ja tehdyt digestiot pipetoitiin geelin kaivoihin. Markkerina käytettiin jälleen Thermo Scientificin GeneRuler 1 kb DNA Ladderia.")
25 Valukelka käännettiin laitteessa ajoasentoon ja geelin päälle kaadettiin valmista 1 x TAE-puskuria, kunnes puskuri peitti geelin täysin. Plasmideille tehtiin digestio ohjeen mukaan (liite 2) ja tehdyt digestiot pipetoitiin geelin kaivoihin. Markkerina käytettiin jälleen Thermo Scientificin GeneRuler 1 kb DNA Ladderia. Ajoohjelmaksi laitteeseen asetettiin 1 tunnin ajoaika ja 300 V tehoa. Ajo pysäytettiin hieman aikaisemmin, koska markkerin väri oli kulkeutunut tarpeeksi alas. Lopuksi geeli kuvattiin UV-kameralla ja siitä tulostettiin kuva 7. Kuvassa näkyy keskimmäisenä markkeri ja reunoilla oikeassa kohdassa olevat vyöhykkeet. KUVA 7. Plasmidien vyöhykkeet ja markkeri agaroosigeelillä 5.4 Sekvensointi Sekvensoinnin tarkoituksena oli selvittää plasmidin sekvenssi ja tarkistaa sen sopivuus COS 7 -soluihin. Sekvensointinäytteet lähetettiin Biocenterin sekvensointilaboratorioon. Näytteet esivalmisteltiin seuraavasti. Normaaliin eppendorfputkeen pipetoitiin 1 µl näytettä, 0,5 µl Forward-primeria ja 6 µl steriiliä ulrapuhdasta vettä. Tulosta verrattiin jo tiedossa olevaan COS 7 -solujen genomiin. Vertailussa todettiin aminohappojärjestyksessä olevan yksi poikkeavuus sekvenssin
.")
26 emäksen kohdalla (kuva 8). KUVA 8. AE2-plasmidin, primerin ja COS 7 -solujen sekvensit 5.5 Transfektio COS 7 -solut transfektoitiin eli solun genomia muokattiin Clone 1 P x plasmidilla. Transfektion aluksi COS 7 -solut jaettiin kuusikuoppalevylle. Seuraavana päivänä tehtiin varsinainen transfektio. PS-putkiin pipetoitiin150 µl puhdasta DMEM:iä (Dulbecco's Modified Eagle Medium). Siihen lisättiin 7 µl FuGene 6:ta eli lipidirakkulaliuosta ja sekoitettiin liuosta pipetin kärkeä putkessa heilutellen. Liuoksen annettiin tasaantua 5 minuuttia. Liuoksen sekaan pipetoitiin 1 µl eli 1,8 µg plasmidia ja liuosta sekoitettiin jälleen pipetinkärkeä heiluttamalla. Liuoksen annettiin seisoa 20 min, jolloin lipidirakkulat sitoutuivat plasmideihin. Tämän jälkeen putkien plasmidiliuos pipetoitiin tiputtelemalla tippoja solujen päälle ympäri kuoppaa. Kuoppalevy siirrettiin mahdollisimman varovasti takaisin CO2-inkubaattoriin kasvamaan + 37 o C:n lämmössä. 5.6 Sorteeraus Sortteeraus eli solujen jakaminen yksittäin kaivoihin kasvamaan suoritettiin Oulun yliopiston Aapistien kampuksella Mikrobiologian laitoksella sijaitsevalla virtaussytometrilla. Solut kuljettiin sinne 6-kuoppalevyllä, jonka ympärille oli kääritty parafilmiä estämään mahdolliset kontaminaatiot. Solut irrotettiin maljalta vasta paikan päällä trypsiinillä ja ne vietiin 15 ml falcon-putkissa virtaussytometrilaitteen luokse. 96-kuppalevyille jaettiin etukäteen 100 µl kasvatuslientä soluja varten. Laite asetettiin jakamaan soluja vihreimmän absorbanssin kohdalta yksi- 24
27 tellen kaivoihin (liite 6). Jokaiseen alareunan viimeisiin kaivoihin laitettiin kuitenkin tulemaan 10 solua kerralla vertailun vuoksi. Aluksi laite jakoi soluja hyvin hitaasti 96-kuoppalevyille, kunnes se tukkeutui täysin. Tässä vaiheessa levyjä oli jaettu jo 6 kpl. Kun tukkeuma saatiin poistettua laitteistosta, jaettiin vielä kaksi levyä kyseisen vihreän kohdan soluja kaivoihin. Solujen absorboimiskohtaa vaihdettiin viimeisen kahden levyn kohdalla. Sortteerauksen jälkeen 96-kuoppalevyt käärittiin parafilmiin kontaminaatioiden estämiseksi ja kuljetettiin solut takaisin Linnanmaan kampuksen soluviljelyyn. Soluviljelyssä 96-kuoppalevyihin lisättiin vielä 100 µl lisää kasvatuslientä ja kuoppalevyt laitettiin varovasti kasvamaan CO2 -inkubaatoriin + 37 o C:seen. 25
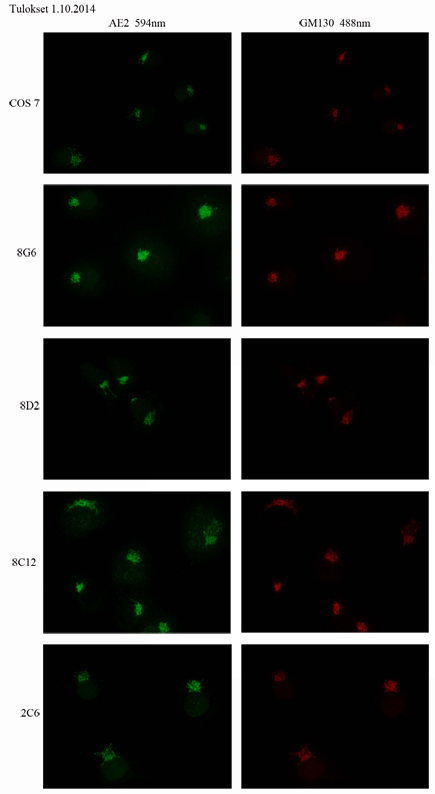
28 6 KLOONIEN KASVATUS JA TARKISTAMINEN 6.1 Kloonien kasvatus Sortteerauksen jälkeen kloonien annettiin olla rauhassa viikon ajan. Sen jälkeen käytiin jokainen 960 kuopasta lävitse ja niistä 12 kaivosta löydettiin jakautuneita soluja. Jokaisesta kaivosta myös poistettiin 100 µl kasvatuslientä ja tilalle laitettiin 150 µl uutta kasvatuslientä. Tuosta liemestä 50 µl oli varattu mahdolliseen haihtumiseen. Viikon päästä jokainen kaivo tarkastettiin jälleen uudelleen ja huomattiin, että muutoksia oli enää 9 kaivossa. Kaikkiin kaivoihin vaihdettiin kuitenkin varalta edellisellä tavalla kasvatusliemi, mahdollisten huomaamattomien kloonien jakautumisen takaamiseksi. Noin kuukauden tarkkailun jälkeen ja kloonien hitaan jakautumisen takia alettiin klooneille laittaa alkuperäisten COS 7 - solujen puhdistettua vanhaa kasvatusliemiseosta. Tässä seoksessa oli puolet kaksi kertaa puhdistettua (kasvatusliemi sentrifugoitiin ja kerättiin ainoastaan varovasti nesteen yläpintaa) COS 7 -soluilla ollutta kasvatuslientä ja puolet uutta puhdasta kasvatuslientä. Tämä sai aikaan sen, että kolme silloisesta 6 kloonista alkoi jakautua paremmin. Viikon kuluttua vanhan kasvatusliemen antamisen jälkeen ensimmäinen 8C12- klooni jaettiin 24-kuoppalevylle. Viikon päästä sen jakamisesta 8C12 jaettiin pienille 3,5 cm:n maljoille, joissa oli pohjalla steriilit peitinlasit. Samalla jakokerralla jaettiin 24-kuoppalevylle uusi klooni 8G6. Neljä päivää sopeutunut 8G6 jaettiin myös pienelle peitinlasilliselle maljalle. Seuraavana päivänä nämä solut värjättiin ensimmäisen kerran, mutta värjäys epäonnistui voimakkaan taustavärin vuoksi. 8G6- ja 8C12-soluja hoidettiin jakamalla niitä isompiin maljoihin ja parin viikon kuluttua jaettiin 8D2-klooni 24-kuoppalevylle. 8D2 jaettiin viikon kuluttua jälleen 6-kuoppalevylle kasvamaan. Tämän jälkeen siirryttiin pitämään soluja vain elossa jakamalla ne säännöllisesti, kunnes värjäysmetodi saatiin toimivaksi. Viimeisenä jaettiin 2C6-solut, jotka olivat olleet hyvin kitukasvuisia koko ajan. Samalla kertaa jaettiin myös 4D5-solut, mutta ne eivät kestäneet jakami- 26
29 sen aiheuttamaa rasitusta ja kuolivat jakamisen yhteydessä. Kun 2C6-solut saatiin kasvamaan tarpeeksi värjäystä varten, jaettiin loputkin 3 kloonia värjäystä varten pienelle peitinlasimaljalle. 6.1 Immonofluoresenssivärjäys Immunofluoresenssivärjäyksen tarkoituksena oli kertoa, oliko soluista onnistuttu poistamaan haluttu geeni. Koska kyseinen geeni vastaa AE2-proteiinin tuotannosta, käytettiin värjäyksessä primaarivasta-aineena AE2-vasta-ainetta. Tällöin pystyttiin näkemään solusta siinä oleva AE2-proteiini. Toisena vasta-aineena käytettiin GM130-vasta-ainetta, joka sitoutuu Golgin laitteessa tuotettavaan toiseen proteiiniin ja näin ollen paljastaa Golgin laitteen sijainnin solussa. Värjäykset tehtiin ohjeen mukaan (liite 3). Immunofluoresenssivärjäyksen kanssa oli alkuun ongelmia taustavärin voimakkuuden takia, ja sen syytä selviteltiin useiden viikon ajan toistuvilla värjäyksillä. Koska värjäykset epäonnistuivat useita kertoja, värjättiin pitkän aikaa vain kotrollisoluina käytettyjä COS 7 -soluja. Näin ollen klooneja tyydyttiin vain jakamaan hengissä selviämisen vuoksi uudelle maljalle. Lopulta syyksi selvisi primaarivasta-aineen liian vähäinen määrä, jolloin solujen normaali fluoresointi tuli värjäyksen läpi. 6.2 Mikroskopia Mikroskooppina käytettiin Olympus BX51 -fluoresenssimikroskooppia. Näytteet kuvattiin aina 60x objektiivilla immersioöljyn kanssa. GM130-vasta-aineen vihreään väriin käytettiin FITC-suodatinta ja AE2-vasta-aineen punaiseen väriin TRITC-suodatinta. Jos solut oli värjätty lisäksi Hoechst-värjäyksellä, ne tarkastettiin DAPI-suodatinta käyttämällä. Jokaisesta vasta-aineesta otettiin kuvat ja niistä tehtiin kuvakollaasi, jotta kuvien vertailu toisiinsa olisi helpompaa. 27
30 7 TULOKSET JA POHDINTA Lopullisia klooneja saatiin selviytymään vain 4 kpl, joista kolme sijaitsi viimeisellä onnistuneella 96-kuoppalevyllä. Immunofluoresenssivärjäyksissä kuitenkin selvisi, ettei mikään klooneista ollut poistogeeninen. Geeni, jota soluista yritettiin poistaa, tuottaa AE2-proteiinia Golgin laitteeseen, ja värjäyksissä keskityttiin todentamaan AE2-proteiinin olemassa olo. Näin ollen poistogeenistäsolulinjaa ei saatu valmistettua. Kuten liitteessä 7 näkyy, GM130- ja AE2-vasta-aineet ovat värjäytyneet täysin samasta kohdasta. Mahdollisia syitä solulinjan valmistuksen epäonnistumiseen on useita. Geeni, joka siirrettiin COS 7 -soluihin, on ihmisperäinen. Kuitenkin solut on alun perin eristetty apinasta, joten geneettinen yhteensopivuus ei ollut paras mahdollinen. Sekvensoinnin tuloksena näkyvässä emäsparijärjestyksessä näkyy yhden emäksen ero oleellisessa kohtaa sekvenssiä. Koska CRISPR-menetelmä on hyvin tarkka ja muuttaa enimmillään vain 20 emäsparin ketjua, on hyvin mahdollista, että kyseinen eroavaisuus on osunut juuri tunnistusalueelle. Jos näin on käynyt, ei soluissa ole tapahtunut halutun AE2-geenin muutosta. Toinen mahdollinen syy epäonnistumiselle oli proteiinin tärkeys solujen elintoiminnoille. Koska kloonien saanto oli hyvin huono, vain 4 kloonia 960 mahdollisuudesta, on tarpeen epäillä, etteivät onnistuneet poistogeeniset kloonit pysyneet hengissä. Tätä teoriaa tukee myös 8 kloonipopulaation kuoleminen heti kasvatuksen alkuvaiheessa. Nämä 8 populaatiota eivät juuri jakautuneet ja näyttivät mikroskoopilla katsottuina hyvin kitukasvuisilta. Kyseisten populaatioiden soluissa oli myös havaittavissa poikkeavuuksia: normaalia suurempaan kokoa, tavallisen soikiomaisen rakenteen sijaan neliskanttisen pitkulaista rakennetta ja solujen jakautumista toistensa päälle vierekkäin levittymisen sijasta. Tämä selittäsi myös, miksi kaikki neljä kloonia olivat vain tavallisia COS 7 -soluja. On myös mahdollista, että selvinneet kloonit ehtivät korjaamaan omaa genomiaan pitkän kasvatusajan puitteissa. Solut pyrkivät aina korjaamaan 28
31 niissä vallitsevat epätasapainot, mutta korjaamisen onnistumiseksi DNA:n toisen juosteen olisi pitänyt jäädä katkeamatta. Vaikka tämä onkin epätodennäköistä, ei vaihtoehtoa voi täysin sulkea pois. Viimeisenä mahdollisena syynä poistogeenisen solulinjan valmistuksen epäonnistumiseen on solujen lajittelu ja sen ongelmat. Lähes kaikki kloonit olivat samalta kuoppalevyltä. Kyseinen levy oli sortteerattu sen jälkeen, kun laitteessa ilmeni tukkeuma. Samainen levy oli myös viimeinen, joka otettiin vielä vihreän absorbanssin alueelta. Tarkkaa syytä työn epäonnistumiselle on kuitenkin mahdotonta antaa. 29
32 8 YHTEENVETO Opinnäytetyön tavoitteena oli valmistaa poistogeeninen solulinja CRISPR-menetelmällä. Solulinja haluttiin valmistaa, jotta olisi voitu tutkia poistetun geenin ja siitä tuotettavan proteiinin vaikutuksia solulle. Menetelmää ei ollut aiemmin kokeiltu tämän solulinjan valmistamiseen. Työn alussa poistogeenistä plasmidia tuotettiin lisää bakteereilla ja valittiin käyttöön sopiva solulinja. Koska tukimusta oli jo aikaisemmin tehty COS 7 -solulinjalla, päätettiin tässäkin työssä käyttää samaista solulinjaa. Lisäksi COS 7 -solut jakautuvat melko nopeasti ja ovat selkeitä värjättyinä. Kasvatuksen ja puhdistamisen jälkeen plasmidi transfektoitiin soluihin. Kun transfektiosta oli kulunut kaksi päivää, solut laskettiin ja jaettiin onnistuneen transfektion solut eli ne, joissa näkyi fluoresenssileima omalle 6-kuoppalevylle. Tämän jälkeen solut vietiin sortteerattaviksi omiin kaivoihinsa 96-kuoppalevyille, joita oli yhteensä 10 kpl. Loppuaika opinnäytetyön teosta oli hidasta solujen jakautumisen odottamista ja niiden elossa pitämistä. Hieman myöhemmin aloitettiin hiomaan immunofluoresenssivärjäysmetodia paremmaksi ja lopulta värjättiin myös klooneja. Klooneja saatiin tuotettua 4 kpl 960 kpl:sta, joista kaikki olivatkin samankaltaisia alkuperäisen solulinjan kanssa. Tämä viittaisi soluista poistetun proteiinin olleen liian tärkeä solun toimintojen kannalta. Koska yksi todennäköinen syy solulinjan valmistamisen epäonnistumiseen oli solulinjan apinaperäisyys ja plasmidin ihmisperäisyys, voisi tätä menetelmää kokeilla uudelleen jossain ihmisperäisessä solulinjassa. Tällöin mahdollinen emäsmuutos tärkeimmän koodausalueen reunoilla ei aiheuttaisi koko solulinjan kuolemista. Hyvä syy eri solulinjan kokeiluun on myös työn alkuvaiheessa tehdyt virheet. Työssä olisi säästetty paljon aikaa, jos heti soluihin trasfektoitumisen jälkeen 30
33 solut olisi sekvensoitu. Tällöin olisi nähty, onko plasmidi oikeasti muokannut genomia eikä vain käynyt hetkellisesti solun sisällä. Näin ollen ei olisi tarvinnut odottaa 6:ta viikkoa ensimmäisen kloonin jakamista. Tätä menetelmää voisi myös kehittää käyttämällä klooneilla vain osittain vanhaa kasvatuslientä, jolloin solukloonit saisivat niille tarpeellisia solujen erittämiä kemiallisia viestejä jakaantumisesta. Jos näin olisi tehty alusta asti, olisi 12 alkuperäisestä kloonista voinut selvitä hengissä useampikin kuin 4 kpl. 31
34 LÄHTEET 1. Aittomäki, Esa Eerikäinen, Tero Leisola, Matti Ojamo, Heikki - Suominen, Ilari Von Weymarn, Niklas Bioprosessitekniikka. Helsinki: WSOY. 2. Solu on toimiva kokonaisuus. Peda.net. Saatavissa: Hakupäivä Solukalvo Solunetti. Saatavissa: Hakupäivä Heino, Jyrki Vuento, Matti Solubiologia. Porvoo: WSOY. 5. Eläinsolu Peda.net. Saatavissa: Hakupäivä Kromatiini Solunetti. Saatavissa: Hakupäivä Clark, Jim DNA structure. Saatavissa: Hakupäivä Haajanen, Kari Pelkonen, Jani Pärssinen, Raimo Suominen, Ilari Geenitekniikka. Turku: Turun ammattikorkeakoulu. 9. Difference DNA RNA Saatavissa: Hakupäivä Tapani, Pentti Elävä solu. Helsinki: Gaudeamus. 11. Aromäki, Kirsi Huuhka, Emma Golgin laite. Saatavissa: Hakupäivä Becker, Wayne M. Bretoni, Gregory Paul Hardin, Jeff Kleinsmith, Lewis J The World of the Cell. 7., painos. San Francisco: Benjamin Cummings. 13. Chatterjee, Titottama Organelles and Their Functions. Saatavissa: Hakupäivä
35 14. Proteiinien tehtäviä Solunetti. Saatavissa: Hakupäivä Proteiinit Solunetti. Saatavissa: Hakupäivä SLC4A2 (Solute Carrier Family-4 Anion Exchanger Member-2). Saatavissa: Hakupäivä Kellokumpu, S. Neff, L. Jämsä-Kellokumpu, S. Kopito, R. Baron, R A 115-kD polypeptide immunologically related to erythrocyte band 3 is present in Golgi membranes. Science vol S Zinc Finger Nuclease (ZFN) Genome-Editing. Saatavissa: Hakupäivä Joung, J. Keith Sander, Jeffry D TALENs: a widely applicable technology for targeted genome editing. Saatavissa: Hakupäivä Bikard, David Cox, David Jiang, Wenyan Marraffini, Luciano A. Zhang, Feng RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature Biotechnology vol. 3. S CRISPR Genome Engineering. Saatavissa: Hakupäivä Plasmid Saatavissa: Hakupäivä Boyer, Rodney Modern experimental biochemistry. 3., painos. San Francisco: Benjamin Cummings. 24. CRISPR (Cas9) Genome-Editing. Horizon. Saatavissa: Hakupäivä Nukleiinihappojen monistamien Solunetti. Saatavissa: Hakupäivä
36 26. Sekvensointi Solunetti. Saatavissa: Hakupäivä Sangerin menetelmä Solunetti. Saatavissa: Hakupäivä Hassinen, Antti Kokkonen, Nina Paakkola, Teija Reini, Kaarina Rivinoja, Antti Cell biology practical part opetusmateriaali. Oulun yliopisto. 29. Flow Cytometry - How Does It Work? Oregon State University. Saatavissa: Hakupäivä Cook, Deborah Odell, Ian D Immunofluorescence Techniques. Journal of Investigative Dermatology vol. 133, nro. 1. Saatavissa: Hakupäivä Vancells, Javier Conde. Direct vs indirect immunofluorescence. Saatavissa: Hakupäivä Soluviljely Solunetti. Saatavissa: Hakupäivä Coté, Rosalie J Aseptic technique for cell culture. Current Protocols in Cell Biology. S Saatavissa: Hakupäivä Phelan, Mary C Basic techniques for mammalian cell tissue culture. Current Protocols in Cell Biology. S Saatavissa: Hakupäivä Animal Cell Culture Protocols & Applications. Qiagen. Saatavissa: Hakupäivä: Solulinjat Solunetti. Saatavissa: Hakupäivä:
37 LIITTEET Liite 1 Työssä käytetyt reagenssit ja laitteisto Liite 2 Transformaatio ja Digestiot Liite 3 Method for Immunofluorescence staining on Coverslips Liite 4 Plasmid DNA Mini Kit työohje Liite 5 Nucleobond Xtra Midi työohje Liite 6 Sortteerauksen solujen valintakohdat. Liite 7 Tulokset 35
38 LIITE 1 TYÖSSÄ KÄYTETYT REAGENSSIT JA LAITTEISTO Reagenssit AE2ct, Teetetty Agaroosi, agarose multi-purpose, Bioline Albumin from bovine serum, Sigma Alexa fluor 488, Invitrogen Alexa fluor 594, Invitrogen Dulbecco's modified eagle's medium (DMEM), Sigma Dulbecco's phosphate buffered saline 10x (PBS), Sigma FuGene 6 transfektion reagent, Promega GeneRuler 1 kb DNA ladder, Thermo Scientific GM130, BD Biosciences Immu-Mount, Thermo Scientific iproof high fidentily DNA polymerase, Bio-Rad Nonident P40, Roche Pensilliini (10 ku/ml streptomysiini (10 mg/ml), Sigma Saponin, Calbiochem Laitteet BIOWIZARD Golden Line, Kojair Centrifuge 5415 D, Eppendorf CO2 Inkubator, Binder Easy-Cast Elektroporesis system Electrophoresis power supply GE Healthcare EPS 301 Electrophoresis Power Supply High performance 2UV transilluminator, UVP Leica DM IL LED microscope Mastercycler personal PCR, Eppendorf Megafuge 1.9 R, Heracus SUB Agua 26, Grant System microscope BX51 BX2 series Vortex-genie
39 LIITE 2 Transformaatio Transformaation of bacterial cell 1. Add DNA (ligation mix 1-2 µg pdna) into 200 µl competent cells. 2. Incubate on ice for 30 min. 3. Heat shock + 42 C, 45 s. (critical step, be accurate wiht the incubation time!) 4. Incubateon ice for 2min after which add 300 µl LB-medium without antibodies. 5. Grow cells 1h in tabletop shaker + 37 C, 300 rpm. 6. Inoculate suspension into LB-plate with proper antibodies (depends on a plasmid) 7. Grow cells overnight in + 37 C. Next day prepare mini-prep cultures if any bacterial colonies exist. Digestiot Tehtiin digestiot PCR_lle seuraavan ohjeen mukaan: PCR-tuote: 25 µl PCR-tuotetta 5 µl 10* restriktioentsyymiin sopivaa puskuria (Fastdigest Green tai Tango) 1 µl BSA:ta (jos tarvitsee, kuten esim. Kpn I) 1 µl 5 entsyymi (10 U) 1 µl 3 entsyymi (yleensä XbaI) (10 U) Inkuboidaan 1-1,5h +37 C:ssa
40 LIITE 3 Ohje immunofluoresenssivärjäykseen Method for Immunofluorescence staining on Coverslips 1. Wash cells twice with 1 ml PBS ph 7.4 (RT) 2. Fix coverslips in 4% paraformaldehyde, 1 ml (diluted in PBS, ph 7.4) for 15 min RT 3. Wash with several changes (3x) PBS (1 ml) 4. Block non-specific staining by incubating in 1 ml PBS + 5% BSA (or FBS) + 0.1% Saponin for 1h at RT. 5. Incubate in primary antibody (100 l; diluted in previous buffer) for 1h at RT. 6. Wash 2 x (1ml) PBS + 5% BSA + 0.1% Saponin 7. Incubate in fluorescent labeled secondary antibody (1: 500; 100 l) for 1h at RT in the dark to protect the fluorescence. 8. a) wash with 2 x PBS + 5% BSA + 0.1% Saponin b) wash in dh2o 9. Mount with Immu-Mount Hoechst staining : Before mounting (and wash with water) incubate the cells with 1 µg/ml Hoechst stain (1 ml) diluted in PBS for 5min. After incubation wash the cells once with PBS and once with water before mounting.
41 LIITE 4/1 E.Z.N.A. Plasmid DNA Mini Kit I Spin Protocol E.Z.N.A. Plasmid DNA Mini Kit I Protocol - Spin Protocol All centrifugation should be performed at room temperature unless otherwise noted. For low copy number plasmids refer to Page 21. This protocol is designed to isolate plasmid DNA from E. coli grown in an overnight 1-5 ml LB culture. Materials and Equipment to be Supplied by User: 100% ethanol Microcentrifuge capable of at least 13,000 x g Nuclease-free 1.5 ml or 2 ml microcentrifuge tubes Culture tubes Optional: sterile deionized water Optional: water bath or incubator capable of 70 C Optional: 3M NaOH solution Before Starting: Heat Elution Buffer to 70 C if plasmid DNA is >10 kb Prepare DNA Wash Buffer and Solution I according to the instructions in the Preparing Reagents section on Page 6 1. Isolate a single colony from a freshly streaked selective plate, and inoculate a culture of 1-5 ml LB medium containing the appropriate selective antibiotic. Incubate for ~12-16 hours at 37 C with vigorous shaking (~ 300 rpm). Use a ml culture tube or a flask with a volume of at least 4 times the volume of the culture. It is strongly recommended that an enda negative strain of E. coli be used for routine plasmid isolation. Examples of such strains include DH5a and JM Centrifuge at 10,000 x g for 1 minute at room temperature. 3. Decant or aspirate and discard the culture media. 4. Add 250 μl Solution I/RNase A. Vortex or pipet up and down to mix thoroughly. Complete resuspension of cell pellet is vital for obtaining good yields. Note: RNase A must be added to Solution I before use. Please see the instructions in the Preparing Reagents section on Page Transfer suspension into a new 1.5 ml microcentrifuge tube. 6. Add 250 μl Solution II. Invert and gently rotate the tube several times to obtain a clear lysate. A 2-3 minute incubation may be necessary. Note: Avoid vigorous mixing as this will shear chromosomal DNA and lower plasmid purity. Do not allow the lysis reaction to proceed more than 5 minutes. Store Solution II tightly capped when not in use to avoid acidification from CO2 in the air. 7. Add 350 μl Solution III. Immediately invert several times until a flocculent white precipitate forms. Note: It is vital that the solution is mixed thoroughly and immediately after the addition of Solution III to avoid localized precipitation. 8. Centrifuge at maximum speed ( 13,000 x g) for 10 minutes. A compact white pellet will form. Promptly proceed to the next step. 9. Insert a HiBind DNA Mini Column into a 2 ml Collection Tube. Optional Protocol for Column Equilibration: 1. Add 100 μl 3M NaOH to the HiBind DNA Mini Column. 2. Centrifuge at maximum speed for seconds. 3. Discard the filtrate and reuse the collection tube. 10. Transfer the cleared supernatant from Step 8 by CAREFULLY aspirating it into the HiBind DNA Mini Column. Be careful not to disturb the pellet and that no cellular debris is transferred to the HiBind DNA Mini Column. 11. Centrifuge at maximum speed for 1 minute. 12. Discard the filtrate and reuse the collection tube. 13. Add 500 μl HB Buffer. 14. Centrifuge at maximum speed for 1 minute.
42 LIITE 4/2 15. Discard the filtrate and reuse collection tube. 16. Add 700 μl DNA Wash Buffer. Note: DNA Wash Buffer must be diluted with 100% ethanol prior to use. Please see Page 6 for instructions. 17. Centrifuge at maximum speed for 1 minute. 18. Discard the filtrate and reuse the collection tube. Optional: Repeat Steps for a second DNA Wash Buffer wash step. 19. Centrifuge the empty HiBind DNA Mini Column for 2 minutes at maximum speed to dry the column matrix. Note: It is important to dry the HiBind DNA Mini Column matrix before elution. Residual ethanol may interfere with downstream applications. 20. Transfer the HiBind DNA Mini Column to a clean 1.5 ml microcentrifuge tube. 21. Add μl Elution Buffer or sterile deionized water directly to the center of the column membrane. Note: The efficiency of eluting DNA from the HiBind DNA Mini Column is dependent on ph. If using sterile deionized water, make sure that the ph is around Let sit at room temperature for 1 minute. 23. Centrifuge at maximum speed for 1 minute. Note: This represents approximately 70% of bound DNA. An optional second elution will yield any residual DNA, though at a lower concentration. 24. Store DNA at -20 C.
43 LIITE 5
44 Sortteerauksen solujen valintakohdat. LIITE 6
45 LIITE 7
2.1 Solun rakenne - Lisämateriaalit
 2.1 Solun rakenne - Lisämateriaalit Tiivistelmä Esitumaisiset eli alkeistumalliset solut ovat pieniä (n.1-10µm), niissä on vähän soluelimiä, eikä tumaa (esim. arkeonit, bakteerit) Tumalliset eli aitotumalliset
2.1 Solun rakenne - Lisämateriaalit Tiivistelmä Esitumaisiset eli alkeistumalliset solut ovat pieniä (n.1-10µm), niissä on vähän soluelimiä, eikä tumaa (esim. arkeonit, bakteerit) Tumalliset eli aitotumalliset
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita
 Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 10. Valkuaisaineiden valmistaminen solussa 1. Avainsanat 2. Perinnöllinen tieto on dna:n emäsjärjestyksessä 3. Proteiinit koostuvat
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 10. Valkuaisaineiden valmistaminen solussa 1. Avainsanat 2. Perinnöllinen tieto on dna:n emäsjärjestyksessä 3. Proteiinit koostuvat
Bioteknologian perustyökaluja
 Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi
 6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
Geenitekniikan perusmenetelmät
 Loppukurssikoe To klo 14-16 2 osiota: monivalintatehtäväosio ja kirjallinen osio, jossa vastataan kahteen kysymykseen viidestä. Koe on auki klo 14.05-16. Voit tehdä sen oppitunnilla, jolloin saat tarvittaessa
Loppukurssikoe To klo 14-16 2 osiota: monivalintatehtäväosio ja kirjallinen osio, jossa vastataan kahteen kysymykseen viidestä. Koe on auki klo 14.05-16. Voit tehdä sen oppitunnilla, jolloin saat tarvittaessa
GEENITEKNIIKAN PERUSASIOITA
 GEENITEKNIIKAN PERUSASIOITA GEENITEKNIIKKKA ON BIOTEKNIIKAN OSA-ALUE! Biotekniikka tutkii ja kehittää elävien solujen, solun osien, biokemiallisten menetelmien sekä molekyylibiologian uusimpien menetelmien
GEENITEKNIIKAN PERUSASIOITA GEENITEKNIIKKKA ON BIOTEKNIIKAN OSA-ALUE! Biotekniikka tutkii ja kehittää elävien solujen, solun osien, biokemiallisten menetelmien sekä molekyylibiologian uusimpien menetelmien
Biopolymeerit. Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä.
 Biopolymeerit Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä. Tärkeimpiä biopolymeerejä ovat hiilihydraatit, proteiinit ja nukleiinihapot. 1 Hiilihydraatit Hiilihydraatit jaetaan mono
Biopolymeerit Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä. Tärkeimpiä biopolymeerejä ovat hiilihydraatit, proteiinit ja nukleiinihapot. 1 Hiilihydraatit Hiilihydraatit jaetaan mono
Metsäpatologian laboratorio tuhotutkimuksen apuna. Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari
 Metsäpatologian laboratorio tuhotutkimuksen apuna Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari Metsäpuiden vaivat Metsäpuiden eloa ja terveyttä uhkaavat monet taudinaiheuttajat: Bioottiset taudinaiheuttajat
Metsäpatologian laboratorio tuhotutkimuksen apuna Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari Metsäpuiden vaivat Metsäpuiden eloa ja terveyttä uhkaavat monet taudinaiheuttajat: Bioottiset taudinaiheuttajat
PCR - tekniikka elintarvikeanalytiikassa
 PCR - tekniikka elintarvikeanalytiikassa Listerian, Salmonellan ja kampylobakteerien tunnistus elintarvikkeista ja rehuista 29.11.2012 Eva Fredriksson-Lidsle Listeria monocytogenes Salmonella (spp) Campylobacter
PCR - tekniikka elintarvikeanalytiikassa Listerian, Salmonellan ja kampylobakteerien tunnistus elintarvikkeista ja rehuista 29.11.2012 Eva Fredriksson-Lidsle Listeria monocytogenes Salmonella (spp) Campylobacter
Genomin ilmentyminen Liisa Kauppi, Genomibiologian tutkimusohjelma
 Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita. BI2 III Perinnöllisyystieteen perusteita 9. Solut lisääntyvät jakautumalla
 Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 9. Solut lisääntyvät jakautumalla 1. Avainsanat 2. Solut lisääntyvät jakautumalla 3. Dna eli deoksiribonukleiinihappo sisältää perimän
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 9. Solut lisääntyvät jakautumalla 1. Avainsanat 2. Solut lisääntyvät jakautumalla 3. Dna eli deoksiribonukleiinihappo sisältää perimän
Solu - perusteet. Enni Kaltiainen
 Solu - perusteet Enni Kaltiainen Solu -perusteet 1. Solusta yleisesti 2. Soluelimet Kalvorakenteet Kalvottomat elimet 3. DNA:n rakenne 4. Solunjakautuminen ja solusykli Synteesi Mitoosi http://www.google.fi/imgres?q=elimet&hl=fi&gbv=2&biw=1280&bih=827&tbm=isch&tbnid=zb_-6_m_rqbtym:&imgrefurl=http://www.hila
Solu - perusteet Enni Kaltiainen Solu -perusteet 1. Solusta yleisesti 2. Soluelimet Kalvorakenteet Kalvottomat elimet 3. DNA:n rakenne 4. Solunjakautuminen ja solusykli Synteesi Mitoosi http://www.google.fi/imgres?q=elimet&hl=fi&gbv=2&biw=1280&bih=827&tbm=isch&tbnid=zb_-6_m_rqbtym:&imgrefurl=http://www.hila
LABORATORIOTYÖ: AGAROOSIGEELIELEKTROFOREESI
 LABORATORIOTYÖ: AGAROOSIGEELIELEKTROFOREESI Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNAjaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Eri valmistajien
LABORATORIOTYÖ: AGAROOSIGEELIELEKTROFOREESI Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNAjaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Eri valmistajien
VASTAUS 1: Yhdistä oikein
 KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
10.9.2015 Pipetointi, sentrifugointi ja spektrofotometria
 BIOKEMIAN MENETELMÄT I, SYKSY 2015 VASTAUKSET LUENTOMATERIAALIN TEHTÄVIIN: 10.9.2015 Pipetointi, sentrifugointi ja spektrofotometria Pesukoneen g-voimat: RCF (x g) = 1,119 10-5 (rpm)2 r = roottorin säde
BIOKEMIAN MENETELMÄT I, SYKSY 2015 VASTAUKSET LUENTOMATERIAALIN TEHTÄVIIN: 10.9.2015 Pipetointi, sentrifugointi ja spektrofotometria Pesukoneen g-voimat: RCF (x g) = 1,119 10-5 (rpm)2 r = roottorin säde
BI4 IHMISEN BIOLOGIA
 BI4 IHMISEN BIOLOGIA IHMINEN ON TOIMIVA KOKONAISUUS Ihmisessä on noin 60 000 miljardia solua Solujen perusrakenne on samanlainen, mutta ne ovat erilaistuneet hoitamaan omia tehtäviään Solujen on oltava
BI4 IHMISEN BIOLOGIA IHMINEN ON TOIMIVA KOKONAISUUS Ihmisessä on noin 60 000 miljardia solua Solujen perusrakenne on samanlainen, mutta ne ovat erilaistuneet hoitamaan omia tehtäviään Solujen on oltava
DNA:n informaation kulku, koostumus
 DNA:n informaation kulku, koostumus KOOSTUMUS Elävien bio-organismien koostumus. Vety, hiili, happi ja typpi muodostavat yli 99% orgaanisten molekyylien rakenneosista. Biomolekyylit voidaan pääosin jakaa
DNA:n informaation kulku, koostumus KOOSTUMUS Elävien bio-organismien koostumus. Vety, hiili, happi ja typpi muodostavat yli 99% orgaanisten molekyylien rakenneosista. Biomolekyylit voidaan pääosin jakaa
Tarkastele kuvaa, muistele matematiikan oppejasi, täytä tekstin aukot ja vastaa kysymyksiin.
 1. Pääryhmien ominaispiirteitä Tarkastele kuvaa, muistele matematiikan oppejasi, täytä tekstin aukot ja vastaa kysymyksiin. Merkitse aukkoihin mittakaavan tuttujen yksiköiden lyhenteet yksiköitä ovat metri,
1. Pääryhmien ominaispiirteitä Tarkastele kuvaa, muistele matematiikan oppejasi, täytä tekstin aukot ja vastaa kysymyksiin. Merkitse aukkoihin mittakaavan tuttujen yksiköiden lyhenteet yksiköitä ovat metri,
PROTEIINIEN MUOKKAUS JA KULJETUS
 PROTEIINIEN MUOKKAUS JA KULJETUS 1.1 Endoplasmakalvosto Endoplasmakalvosto on organelli joka sijaitsee tumakalvossa kiinni. Se on topologisesti siis yhtä tumakotelon kanssa. Se koostuu kahdesta osasta:
PROTEIINIEN MUOKKAUS JA KULJETUS 1.1 Endoplasmakalvosto Endoplasmakalvosto on organelli joka sijaitsee tumakalvossa kiinni. Se on topologisesti siis yhtä tumakotelon kanssa. Se koostuu kahdesta osasta:
Solun perusrakenne I Solun perusrakenne. BI2 I Solun perusrakenne 2. Solun perusrakenne
 Solun perusrakenne I Solun perusrakenne 2. Solun perusrakenne 1. Avainsanat 2. Kaikille soluille yhteiset piirteet 3. Kasvisolun rakenne 4. Eläinsolun rakenne 5. Sienisolun rakenne 6. Bakteerisolun rakenne
Solun perusrakenne I Solun perusrakenne 2. Solun perusrakenne 1. Avainsanat 2. Kaikille soluille yhteiset piirteet 3. Kasvisolun rakenne 4. Eläinsolun rakenne 5. Sienisolun rakenne 6. Bakteerisolun rakenne
GMO analytiikka Annikki Welling Kemian tutkimusyksikkö Evira
 GMO analytiikka Annikki Welling Kemian tutkimusyksikkö Evira Millaisia GM kasvit ovat ja kuinka tätä käytetään hyväksi analytiikassa Aromaattisten aminohappojen biosynteesireitti kasvissa Kasvi tarvitsee
GMO analytiikka Annikki Welling Kemian tutkimusyksikkö Evira Millaisia GM kasvit ovat ja kuinka tätä käytetään hyväksi analytiikassa Aromaattisten aminohappojen biosynteesireitti kasvissa Kasvi tarvitsee
Plasmidi-DNA:n eristys bakteerisoluista DNA:n geelielektroforeesi (Proteiinien geelielektroforeesi)
") Plasmidi-DNA:n eristys bakteerisoluista DNA:n geelielektroforeesi (Proteiinien geelielektroforeesi) CHEM-A1310 Biotieteen perusteet Heli Viskari 2017 DNA-harjoitustöiden aikataulu, valitse yksi näistä
Plasmidi-DNA:n eristys bakteerisoluista DNA:n geelielektroforeesi (Proteiinien geelielektroforeesi) CHEM-A1310 Biotieteen perusteet Heli Viskari 2017 DNA-harjoitustöiden aikataulu, valitse yksi näistä
NaturaPura Ibérica Elokuu 10, 2009 Rua das Australias, No. 1 4705-322 Braga Portugali
 NaturaPura Ibérica Elokuu 10, 2009 Rua das Australias, No. 1 4705-322 Braga Portugali VIITATEN: IN VITRO IHOÄRSYTTÄVYYSTESTAUSRAPORTTI Oheisena NaturaPuran toimittaman 100% puuvillakangasmateriaalin in
NaturaPura Ibérica Elokuu 10, 2009 Rua das Australias, No. 1 4705-322 Braga Portugali VIITATEN: IN VITRO IHOÄRSYTTÄVYYSTESTAUSRAPORTTI Oheisena NaturaPuran toimittaman 100% puuvillakangasmateriaalin in
Synteettinen biologia Suomessa: Virukset synteettisen biologian työkaluina
 Synteettinen biologia Suomessa: Virukset synteettisen biologian työkaluina Minna Poranen Akatemiatutkija Helsingin yliopisto FinSynBio-ohjelma Suomen Akatemia Virukset synteettisen biologian työkaluina
Synteettinen biologia Suomessa: Virukset synteettisen biologian työkaluina Minna Poranen Akatemiatutkija Helsingin yliopisto FinSynBio-ohjelma Suomen Akatemia Virukset synteettisen biologian työkaluina
Biologian tehtävien vastaukset ja selitykset
 Biologian tehtävien vastaukset ja selitykset Ilmainen lääkiksen harjoituspääsykoe, kevät 2017 Tehtävä 2. (20 p) A. 1. EPÄTOSI. Ks. s. 4. Menetelmää käytetään geenitekniikassa geenien muokkaamisessa. 2.
Biologian tehtävien vastaukset ja selitykset Ilmainen lääkiksen harjoituspääsykoe, kevät 2017 Tehtävä 2. (20 p) A. 1. EPÄTOSI. Ks. s. 4. Menetelmää käytetään geenitekniikassa geenien muokkaamisessa. 2.
LABORATORIOTYÖ: RESTRIKTIOENTSYYMIDIGESTIO
 LABORATORIOTYÖ: RESTRIKTIOENTSYYMIDIGESTIO Restriktioentsyymidigestio on yksi yhdistelmä-dna-tekniikan perusmenetelmistä, jossa katkaistaan kaksinauhainen DNA tietystä kohdasta erilaisten restriktioentsyymien
LABORATORIOTYÖ: RESTRIKTIOENTSYYMIDIGESTIO Restriktioentsyymidigestio on yksi yhdistelmä-dna-tekniikan perusmenetelmistä, jossa katkaistaan kaksinauhainen DNA tietystä kohdasta erilaisten restriktioentsyymien
Genetiikan perusteiden harjoitustyöt
 Genetiikan perusteiden harjoitustyöt Molekyylien kloonaus ja siihen liittyvät taidot ja temput, osa 1 Restriktioentsyymit, elektroforeesi Moniste sivulta 24-: Geenien kloonaus CELL 491- Isolating, cloning,
Genetiikan perusteiden harjoitustyöt Molekyylien kloonaus ja siihen liittyvät taidot ja temput, osa 1 Restriktioentsyymit, elektroforeesi Moniste sivulta 24-: Geenien kloonaus CELL 491- Isolating, cloning,
Biomolekyylit ja biomeerit
 Biomolekyylit ja biomeerit Polymeerit ovat hyvin suurikokoisia, pitkäketjuisia molekyylejä, jotka muodostuvat monomeereista joko polyadditio- tai polykondensaatioreaktiolla. Polymeerit Synteettiset polymeerit
Biomolekyylit ja biomeerit Polymeerit ovat hyvin suurikokoisia, pitkäketjuisia molekyylejä, jotka muodostuvat monomeereista joko polyadditio- tai polykondensaatioreaktiolla. Polymeerit Synteettiset polymeerit
Etunimi: Henkilötunnus:
 Kokonaispisteet: Lue oheinen artikkeli ja vastaa kysymyksiin 1-25. Huomaa, että artikkelista ei löydy suoraan vastausta kaikkiin kysymyksiin, vaan sinun tulee myös tuntea ja selittää tarkemmin artikkelissa
Kokonaispisteet: Lue oheinen artikkeli ja vastaa kysymyksiin 1-25. Huomaa, että artikkelista ei löydy suoraan vastausta kaikkiin kysymyksiin, vaan sinun tulee myös tuntea ja selittää tarkemmin artikkelissa
Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20
 Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20 3: Osa 1 Tumallisten solujen genomin toiminnassa sekä geenien
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20 3: Osa 1 Tumallisten solujen genomin toiminnassa sekä geenien
DNA, RNA ja proteiinirakenteen ennustaminen
 S-114.500 Solubiosysteemien perusteet Harjoitustyö Syksy 2003 DNA, RNA ja proteiinirakenteen ennustaminen Ilpo Tertsonen, 58152p Jaakko Niemi, 55114s Sisällysluettelo 1. Alkusanat... 3 2. Johdanto... 4
S-114.500 Solubiosysteemien perusteet Harjoitustyö Syksy 2003 DNA, RNA ja proteiinirakenteen ennustaminen Ilpo Tertsonen, 58152p Jaakko Niemi, 55114s Sisällysluettelo 1. Alkusanat... 3 2. Johdanto... 4
"Geenin toiminnan säätely" Moniste sivu 13
 "Geenin toiminnan säätely" Moniste sivu 13 Monisteen alussa on erittäin tärkeitä ohjeita turvallisuudesta Lukekaa sivu 5 huolellisesti ja usein Vaarat vaanivat: Palavia nesteitä ja liekkejä on joskus/usein
"Geenin toiminnan säätely" Moniste sivu 13 Monisteen alussa on erittäin tärkeitä ohjeita turvallisuudesta Lukekaa sivu 5 huolellisesti ja usein Vaarat vaanivat: Palavia nesteitä ja liekkejä on joskus/usein
Mitä elämä on? Astrobiologian luento 15.9.2015 Kirsi
 Mitä elämä on? Astrobiologian luento 15.9.2015 Kirsi Määritelmän etsimistä Lukemisto: Origins of Life and Evolution of the Biosphere, 2010, issue 2., selaile kokonaan Perintteisesti: vaikeasti määriteltävä
Mitä elämä on? Astrobiologian luento 15.9.2015 Kirsi Määritelmän etsimistä Lukemisto: Origins of Life and Evolution of the Biosphere, 2010, issue 2., selaile kokonaan Perintteisesti: vaikeasti määriteltävä
Anatomia ja fysiologia 1 Peruselintoiminnat
 Anatomia ja fysiologia 1 Peruselintoiminnat Solu Laura Partanen Yleistä Elimistö koostuu soluista ja soluväliaineesta Makroskooppinen mikroskooppinen Mm. liikkumiskyky, reagointi ärsykkeisiin, aineenvaihdunta
Anatomia ja fysiologia 1 Peruselintoiminnat Solu Laura Partanen Yleistä Elimistö koostuu soluista ja soluväliaineesta Makroskooppinen mikroskooppinen Mm. liikkumiskyky, reagointi ärsykkeisiin, aineenvaihdunta
Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe Tehtävä 1 Pisteet / 15
 Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
Solun perusrakenne I Solun perusrakenne. BI2 I Solun perusrakenne 3. Solujen kemiallinen rakenne
 Solun perusrakenne I Solun perusrakenne 3. Solujen kemiallinen rakenne 1. Avainsanat 2. Solut koostuvat molekyyleistä 3. Hiilihydraatit 4. Lipidit eli rasva-aineet 5. Valkuaisaineet eli proteiinit rakentuvat
Solun perusrakenne I Solun perusrakenne 3. Solujen kemiallinen rakenne 1. Avainsanat 2. Solut koostuvat molekyyleistä 3. Hiilihydraatit 4. Lipidit eli rasva-aineet 5. Valkuaisaineet eli proteiinit rakentuvat
Henkilötunnus - Biokemian/bioteknologian valintakoe. Sukunimi Etunimet Tehtävä 1 Pisteet / 20
 elsingin yliopisto/tampereen yliopisto enkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24. 5. 2004 Etunimet Tehtävä 1 Pisteet / 20 Solujen kalvorakenteet rajaavat solut niiden ulkoisesta ympäristöstä
elsingin yliopisto/tampereen yliopisto enkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24. 5. 2004 Etunimet Tehtävä 1 Pisteet / 20 Solujen kalvorakenteet rajaavat solut niiden ulkoisesta ympäristöstä
E. colin auksotrofiset mutantit
 E. colin auksotrofiset mutantit Mutantit MCB 261 -; CELL 267 Mutaatio tapahtuu DNA:ssa, mutta vaikutus näkyy tai tuntuu vasta jos proteiini muuttuu oleellisesti Auksotrofinen mutantti tarkoittaa sellaista
E. colin auksotrofiset mutantit Mutantit MCB 261 -; CELL 267 Mutaatio tapahtuu DNA:ssa, mutta vaikutus näkyy tai tuntuu vasta jos proteiini muuttuu oleellisesti Auksotrofinen mutantti tarkoittaa sellaista
Genomin ylläpito Tiina Immonen BLL Lääke8eteellinen biokemia ja kehitysbiologia
 Genomin ylläpito 14.1.2014 Tiina Immonen BLL Lääke8eteellinen biokemia ja kehitysbiologia Luennon sisältö DNA:n kahdentuminen eli replikaa8o DNA:n korjausmekanismit Replikaa8ovirheiden korjaus Emäksenpoistokorjaus
Genomin ylläpito 14.1.2014 Tiina Immonen BLL Lääke8eteellinen biokemia ja kehitysbiologia Luennon sisältö DNA:n kahdentuminen eli replikaa8o DNA:n korjausmekanismit Replikaa8ovirheiden korjaus Emäksenpoistokorjaus
Vanilliini (karbonyyliyhdiste) Etikkahappo (karboksyyliyhdiste)
 Etikkahappo (karboksyyliyhdiste)") 1 a) Määrittele karbonyyliyhdiste. Piirrä esimerkkirakennekaava ja nimeä se. Samoin määrittele karboksyyliyhdiste, piirrä esimerkkirakennekaava ja nimeä se. Toisen esimerkin tulee olla rakenteeltaan avoketjuinen,
1 a) Määrittele karbonyyliyhdiste. Piirrä esimerkkirakennekaava ja nimeä se. Samoin määrittele karboksyyliyhdiste, piirrä esimerkkirakennekaava ja nimeä se. Toisen esimerkin tulee olla rakenteeltaan avoketjuinen,
Esim. ihminen koostuu 3,72 x solusta
 Esim. ihminen koostuu 3,72 x 10 13 solusta Erilaisia soluja Veren punasoluja Tohvelieläin koostuu vain yhdestä solusta Siittiösolu on ihmisen pienimpiä soluja Pajun juurisolukko Bakteereja Malarialoisioita
Esim. ihminen koostuu 3,72 x 10 13 solusta Erilaisia soluja Veren punasoluja Tohvelieläin koostuu vain yhdestä solusta Siittiösolu on ihmisen pienimpiä soluja Pajun juurisolukko Bakteereja Malarialoisioita
- Extra: PCR-alukkeiden suunnittelutehtävä haluttaessa
 Kertaus CHEM-C2300 0 Tällä luennolla: - Oletteko lukeneet artikkelia, käydäänkö läpi? - Ehdotuksia tenttikysymyksiin? - Käydään läpi kurssin keskeiset asiakokonaisuudet otsikkotasolla - Extra: PCR-alukkeiden
Kertaus CHEM-C2300 0 Tällä luennolla: - Oletteko lukeneet artikkelia, käydäänkö läpi? - Ehdotuksia tenttikysymyksiin? - Käydään läpi kurssin keskeiset asiakokonaisuudet otsikkotasolla - Extra: PCR-alukkeiden
VALINTAKOE 2014 Terveyden biotieteiden koulutusohjelmat/ty ja ISY
 VALINTAKOE 2014 Terveyden biotieteiden koulutusohjelmat/ty ja ISY BIOLOGIAN KYSYMYSTEN Hyvän vastauksen piirteet 2014 Väittämätehtävät. Maksimipisteet 10. Määrittele tai kuvaa lyhyesti seuraavat termit.
VALINTAKOE 2014 Terveyden biotieteiden koulutusohjelmat/ty ja ISY BIOLOGIAN KYSYMYSTEN Hyvän vastauksen piirteet 2014 Väittämätehtävät. Maksimipisteet 10. Määrittele tai kuvaa lyhyesti seuraavat termit.
Bioteknologian tutkinto-ohjelma Valintakoe Tehtävä 3 Pisteet / 30
 Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 3 Pisteet / 30 3. a) Alla on lyhyt jakso dsdna:ta, joka koodaa muutaman aminohappotähteen
Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 3 Pisteet / 30 3. a) Alla on lyhyt jakso dsdna:ta, joka koodaa muutaman aminohappotähteen
9/30/2013. GMO analytiikka. Termistöä. Markkinoilla olevien GM kasvien ominaisuuksia
 GMO analytiikka Kemian ja toksikologian tutkimusyksikkö Evira Termistöä geenimuuntelu muuntogeeninen siirtogeeninen GM GMO (geneettisesti muunnettu organismi) GM tapahtuma (event): käytetään silloin kun
GMO analytiikka Kemian ja toksikologian tutkimusyksikkö Evira Termistöä geenimuuntelu muuntogeeninen siirtogeeninen GM GMO (geneettisesti muunnettu organismi) GM tapahtuma (event): käytetään silloin kun
Tarja Huhta. FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA
 Tarja Huhta FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA Tarja Huhta Opinnäytetyö Kevät 2012 Laboratorioalan koulutusohjelma Oulun
Tarja Huhta FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA Tarja Huhta Opinnäytetyö Kevät 2012 Laboratorioalan koulutusohjelma Oulun
TEE-SE-ITSE-ELEKTROFOREESI
 TEE-SE-ITSE-ELEKTROFOREESI Justus Mutanen (Opinkirjon työn pohjalta) BioPop-resurssikeskus, Helsingin yliopiston LUMA-keskus Työn tavoite Työn tavoitteena on tutustua elektroforeesin periaatteeseen ja
TEE-SE-ITSE-ELEKTROFOREESI Justus Mutanen (Opinkirjon työn pohjalta) BioPop-resurssikeskus, Helsingin yliopiston LUMA-keskus Työn tavoite Työn tavoitteena on tutustua elektroforeesin periaatteeseen ja
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. a) Seoksen komponentit voidaan erotella toisistaan kromatografisilla menetelmillä. Mihin kromatografiset menetelmät perustuvat? (2p) Menetelmät perustuvat seoksen osasten erilaiseen sitoutumiseen paikallaan
1. a) Seoksen komponentit voidaan erotella toisistaan kromatografisilla menetelmillä. Mihin kromatografiset menetelmät perustuvat? (2p) Menetelmät perustuvat seoksen osasten erilaiseen sitoutumiseen paikallaan
måndag 10 februari 14 Jaana Ohtonen Kielikoulu/Språkskolan Haparanda
 GENETIIKKA: KROMOSOMI DNA & GEENI Yksilön ominaisuudet 2 Yksilön ominaisuudet Perintötekijät 2 Yksilön ominaisuudet Perintötekijät Ympäristötekijät 2 Perittyjä ominaisuuksia 3 Leukakuoppa Perittyjä ominaisuuksia
GENETIIKKA: KROMOSOMI DNA & GEENI Yksilön ominaisuudet 2 Yksilön ominaisuudet Perintötekijät 2 Yksilön ominaisuudet Perintötekijät Ympäristötekijät 2 Perittyjä ominaisuuksia 3 Leukakuoppa Perittyjä ominaisuuksia
DNA (deoksiribonukleiinihappo)
") DNA (deoksiribonukleiinihappo) Kaksoiskierre (10 emäsparin välein täysi kierros) Kaksi sokerifosfaattirunkoa. Huomaa suunta: 5 -päässä vapaana fosfaatti (kiinni sokerin 5. hiilessä) 3 -päässä vapaana sokeri
DNA (deoksiribonukleiinihappo) Kaksoiskierre (10 emäsparin välein täysi kierros) Kaksi sokerifosfaattirunkoa. Huomaa suunta: 5 -päässä vapaana fosfaatti (kiinni sokerin 5. hiilessä) 3 -päässä vapaana sokeri
DNA Tiina Immonen, FT, yo-lehtori HY Biolääketieteen laitos, Biokemia ja kehitysbiologia
 DNA 3.3.2015 Tiina Immonen, FT, yo-lehtori HY Biolääketieteen laitos, Biokemia ja kehitysbiologia Koordinaattori, Master s Degree Programme in Translational Medicine (TRANSMED) 1 Sisältö DNA:n rakenne
DNA 3.3.2015 Tiina Immonen, FT, yo-lehtori HY Biolääketieteen laitos, Biokemia ja kehitysbiologia Koordinaattori, Master s Degree Programme in Translational Medicine (TRANSMED) 1 Sisältö DNA:n rakenne
Geenisakset (CRISPR)- Geeniterapian vallankumousko? BMOL Juha Partanen
- Geeniterapian vallankumousko? BMOL Juha Partanen") Geenisakset (CRISPR)- Geeniterapian vallankumousko? BMOL 19.11.2016 Juha Partanen Geenisakset 2 2 N A T U R E V O L 5 2 2 4 J U N E 2 0 1 5 Sisältö Geenimuokkaus: historiallinen perspektiivi Geenisakset
Geenisakset (CRISPR)- Geeniterapian vallankumousko? BMOL 19.11.2016 Juha Partanen Geenisakset 2 2 N A T U R E V O L 5 2 2 4 J U N E 2 0 1 5 Sisältö Geenimuokkaus: historiallinen perspektiivi Geenisakset
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa
 Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa Maria Valkonen, Kaisa Jalkanen, Martin Täubel, Anne Hyvärinen 31.3.2014 Sisäilmastoseminaari 2014 1 Tausta Asumisterveysoppaan mukaiset sisäympäristön
Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa Maria Valkonen, Kaisa Jalkanen, Martin Täubel, Anne Hyvärinen 31.3.2014 Sisäilmastoseminaari 2014 1 Tausta Asumisterveysoppaan mukaiset sisäympäristön
Elämän synty. Matti Leisola
 Elämän synty Matti Leisola Selitettävää Universumin rakenne Biologinen elämä Maailmallemme on olemassa kaksi erilaista selitysmallia Kaikki on syntynyt sattumanvaraisten fysikaalisten ja kemiallisten tapahtumien
Elämän synty Matti Leisola Selitettävää Universumin rakenne Biologinen elämä Maailmallemme on olemassa kaksi erilaista selitysmallia Kaikki on syntynyt sattumanvaraisten fysikaalisten ja kemiallisten tapahtumien
Francis Crick ja James D. Watson
 Francis Crick ja James D. Watson Francis Crick ja James D. Watson selvittivät DNAn rakenteen 1953 (Nobel-palkinto 1962). Rosalind Franklin ei ehtinyt saada kunniaa DNA:n rakenteen selvittämisestä. Hän
Francis Crick ja James D. Watson Francis Crick ja James D. Watson selvittivät DNAn rakenteen 1953 (Nobel-palkinto 1962). Rosalind Franklin ei ehtinyt saada kunniaa DNA:n rakenteen selvittämisestä. Hän
Original Elche antimicrobi TM desinfiointiaineen testaus Legionella lajeille
 Original Elche antimicrobi TM desinfiointiaineen testaus Legionella lajeille Tutkimusraportti 145606 10.3.2006 Sivu 1:6 Sisällysluettelo 1. YHTEYSTIEDOT... 3 2. TESTATTAVAT LEGIONELLA-LAJIT... 3 3. TESTAUSMENETELMÄT...
Original Elche antimicrobi TM desinfiointiaineen testaus Legionella lajeille Tutkimusraportti 145606 10.3.2006 Sivu 1:6 Sisällysluettelo 1. YHTEYSTIEDOT... 3 2. TESTATTAVAT LEGIONELLA-LAJIT... 3 3. TESTAUSMENETELMÄT...
Entsyymit ja niiden tuotanto. Niklas von Weymarn, VTT Erikoistutkija ja tiiminvetäjä
 Entsyymit ja niiden tuotanto Niklas von Weymarn, VTT Erikoistutkija ja tiiminvetäjä Mitä ovat entsyymit? Entsyymit ovat proteiineja (eli valkuaisaineita), jotka vauhdittavat (katalysoivat) kemiallisia
Entsyymit ja niiden tuotanto Niklas von Weymarn, VTT Erikoistutkija ja tiiminvetäjä Mitä ovat entsyymit? Entsyymit ovat proteiineja (eli valkuaisaineita), jotka vauhdittavat (katalysoivat) kemiallisia
Solun toiminta. II Solun toiminta. BI2 II Solun toiminta 8. Solut tarvitsevat energiaa
 Solun toiminta II Solun toiminta 8. Solut tarvitsevat energiaa 1. Avainsanat 2. Solut tarvitsevat jatkuvasti energiaa 3. Soluhengitys 4. Käymisreaktiot 5. Auringosta ATP:ksi 6. Tehtävät 7. Kuvat Avainsanat:
Solun toiminta II Solun toiminta 8. Solut tarvitsevat energiaa 1. Avainsanat 2. Solut tarvitsevat jatkuvasti energiaa 3. Soluhengitys 4. Käymisreaktiot 5. Auringosta ATP:ksi 6. Tehtävät 7. Kuvat Avainsanat:
Peptidi ---- F ----- K ----- V ----- R ----- H ----- A ---- A. Siirtäjä-RNA:n (trna:n) (3 ) AAG UUC CAC GCA GUG CGU (5 ) antikodonit
 (3 ) AAG UUC CAC GCA GUG CGU (5 ) antikodonit") Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 3 Pisteet / 20 Osa 1: Haluat selvittää -- F -- K -- V -- R -- H -- A peptidiä
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 3 Pisteet / 20 Osa 1: Haluat selvittää -- F -- K -- V -- R -- H -- A peptidiä
Euromit2014-konferenssin tausta-aineistoa Tuottaja Tampereen yliopiston viestintä
 Mitkä mitokondriot? Lyhyt johdatus geenitutkijoiden maailmaan Ihmisen kasvua ja kehitystä ohjaava informaatio on solun tumassa, DNA:ssa, josta se erilaisten prosessien kautta päätyy ohjaamaan elimistön,
Mitkä mitokondriot? Lyhyt johdatus geenitutkijoiden maailmaan Ihmisen kasvua ja kehitystä ohjaava informaatio on solun tumassa, DNA:ssa, josta se erilaisten prosessien kautta päätyy ohjaamaan elimistön,
Ekologiset ympäristöongelmat. 10. Geeniteknologia. BI5 II Geeniteknologia 4. Geenitekniikan perusmenetelmiä
 Ekologiset ympäristöongelmat 10. Geeniteknologia Dna:n ja rna:n käsittely Eristäminen Puhdistaminen Lähetti-rna:t voidaan muuntaa niiden emäsjärjestystä vastaavaksi ns. komplementaariseksi dna:ksi (c-dna)
Ekologiset ympäristöongelmat 10. Geeniteknologia Dna:n ja rna:n käsittely Eristäminen Puhdistaminen Lähetti-rna:t voidaan muuntaa niiden emäsjärjestystä vastaavaksi ns. komplementaariseksi dna:ksi (c-dna)
KOE 6 Biotekniikka. 1. Geenien kloonaus plasmidien avulla.
 Esseekysymyksistä 1-2 voi saada enintään 9 pistettä/kysymys. Vastauksia pisteytettäessä huomioidaan asiatiedot, joista voi saada enintään 7 pistettä. Lisäksi vastaaja saa enintään kaksi pistettä, mikäli
Esseekysymyksistä 1-2 voi saada enintään 9 pistettä/kysymys. Vastauksia pisteytettäessä huomioidaan asiatiedot, joista voi saada enintään 7 pistettä. Lisäksi vastaaja saa enintään kaksi pistettä, mikäli
COXSACKIEVIRUS A9 KANTOJEN MUUNTUMINEN
 Opinnäytetyö (AMK) Bio ja elintarviketekniikka Biotekniikka 2011 Tiina Kelanne COXSACKIEVIRUS A9 KANTOJEN MUUNTUMINEN VP1- ja 3D-geenien sekvensointi rekombinaatioiden tunnistamiseksi OPINNÄYTETYÖ (AMK)
Opinnäytetyö (AMK) Bio ja elintarviketekniikka Biotekniikka 2011 Tiina Kelanne COXSACKIEVIRUS A9 KANTOJEN MUUNTUMINEN VP1- ja 3D-geenien sekvensointi rekombinaatioiden tunnistamiseksi OPINNÄYTETYÖ (AMK)
MYKOPLASMA- WORKSHOP!
 Science whereyou are Tarjoukset voimassa 31.12.2014 asti kampanjakoodilla FIN311214FIN. Kampanjahinnat ilman arvonlisäveroa. TULOSSA... MYKOPLASMA- WORKSHOP! BioNordika Finland Oy Kutomotie 18, 00380 Helsinki
Science whereyou are Tarjoukset voimassa 31.12.2014 asti kampanjakoodilla FIN311214FIN. Kampanjahinnat ilman arvonlisäveroa. TULOSSA... MYKOPLASMA- WORKSHOP! BioNordika Finland Oy Kutomotie 18, 00380 Helsinki
Genomin ilmentyminen
 Kauppi 17/01/2014 Genomin ilmentyminen LH1, Molekyylibiologia 17.1.2014 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Huone C501b, Biomedicum 1 Transkriptiofaktorin mutaatio voi
Kauppi 17/01/2014 Genomin ilmentyminen LH1, Molekyylibiologia 17.1.2014 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Huone C501b, Biomedicum 1 Transkriptiofaktorin mutaatio voi
NON-CODING RNA (ncrna)
") NON-CODING RNA (ncrna) 1. Yleistä NcRNA eli non-coding RNA tarkoittaa kaikkia proteiinia koodaamattomia rnamolekyylejä. Näistä yleisimmin tunnetut ovat ribosomaalinen RNA (rrna) sekä siirtäjä-rna (trna),
NON-CODING RNA (ncrna) 1. Yleistä NcRNA eli non-coding RNA tarkoittaa kaikkia proteiinia koodaamattomia rnamolekyylejä. Näistä yleisimmin tunnetut ovat ribosomaalinen RNA (rrna) sekä siirtäjä-rna (trna),
Perinnöllisyyden perusteita
 Perinnöllisyyden perusteita Eero Lukkari Tämä artikkeli kertoo perinnöllisyyden perusmekanismeista johdantona muille jalostus- ja terveysaiheisille artikkeleille. Koirien, kuten muidenkin eliöiden, perimä
Perinnöllisyyden perusteita Eero Lukkari Tämä artikkeli kertoo perinnöllisyyden perusmekanismeista johdantona muille jalostus- ja terveysaiheisille artikkeleille. Koirien, kuten muidenkin eliöiden, perimä
Biomolekyylit 2. Nukleotidit, aminohapot ja proteiinit
 Biomolekyylit 2 Nukleotidit, aminohapot ja proteiinit Nukleotidit Ihmisen perimä, eli DNA (deoksiribonukleiinihappo) muodostuu pitkästä nukleotidiketjusta. Lisäksi nukleotidit toimivat mm. proteiinisynteesissä
Biomolekyylit 2 Nukleotidit, aminohapot ja proteiinit Nukleotidit Ihmisen perimä, eli DNA (deoksiribonukleiinihappo) muodostuu pitkästä nukleotidiketjusta. Lisäksi nukleotidit toimivat mm. proteiinisynteesissä
Nimi sosiaaliturvatunnus
 Valintakoe 2013 / Biokemia Nimi sosiaaliturvatunnus 1. Selitä: (3,0 p) a) Mitä ovat eksonit ja intronit ja miten ne eroavat toisistaan? b) Mitä eläinsolulle tapahtuu, jos se laitetaan sen sisällä olevaa
Valintakoe 2013 / Biokemia Nimi sosiaaliturvatunnus 1. Selitä: (3,0 p) a) Mitä ovat eksonit ja intronit ja miten ne eroavat toisistaan? b) Mitä eläinsolulle tapahtuu, jos se laitetaan sen sisällä olevaa
Biologia. Pakolliset kurssit. 1. Eliömaailma (BI1)
") Biologia Pakolliset kurssit 1. Eliömaailma (BI1) tuntee elämän tunnusmerkit ja perusedellytykset sekä tietää, miten elämän ilmiöitä tutkitaan ymmärtää, mitä luonnon monimuotoisuus biosysteemien eri tasoilla
Biologia Pakolliset kurssit 1. Eliömaailma (BI1) tuntee elämän tunnusmerkit ja perusedellytykset sekä tietää, miten elämän ilmiöitä tutkitaan ymmärtää, mitä luonnon monimuotoisuus biosysteemien eri tasoilla
Solun tutkiminen. - Geenitekniikka
 Solun tutkiminen - Geenitekniikka Tunnin sisältö 1. Bioteknologian peruskäsitteitä 2. Hieman mikroskoopeista 3. DNA:n eristäminen, puhdistaminen ja pilkkominen 4. Geenikirjasto 5. PCR 6. Elektroforeesi
Solun tutkiminen - Geenitekniikka Tunnin sisältö 1. Bioteknologian peruskäsitteitä 2. Hieman mikroskoopeista 3. DNA:n eristäminen, puhdistaminen ja pilkkominen 4. Geenikirjasto 5. PCR 6. Elektroforeesi
DNA (deoksiribonukleiinihappo)
") DNA (deoksiribonukleiinihappo) Kaksoiskierre (10 emäsparin välein täysi kierros) Kaksi sokerifosfaattirunkoa. Huomaa suunta: 5 päässä vapaana fosfaatti (kiinni sokerin 5. hiilessä) 3 päässä vapaana sokeri
DNA (deoksiribonukleiinihappo) Kaksoiskierre (10 emäsparin välein täysi kierros) Kaksi sokerifosfaattirunkoa. Huomaa suunta: 5 päässä vapaana fosfaatti (kiinni sokerin 5. hiilessä) 3 päässä vapaana sokeri
Nukleiinihapot! Juha Klefström, Biolääketieteen laitos/biokemia ja genomibiologian tutkimusohjelma Helsingin yliopisto.
 Nukleiinihapot! Juha Klefström, Biolääketieteen laitos/biokemia ja genomibiologian tutkimusohjelma Helsingin yliopisto Juha.Klefstrom@helsinki.fi Nukleiinihapot! kertausta matkan varrella, vähemmän kuitenkin
Nukleiinihapot! Juha Klefström, Biolääketieteen laitos/biokemia ja genomibiologian tutkimusohjelma Helsingin yliopisto Juha.Klefstrom@helsinki.fi Nukleiinihapot! kertausta matkan varrella, vähemmän kuitenkin
Vinkkejä opettajille ja odotetut tulokset SIVU 1
 Vinkkejä opettajille ja odotetut tulokset SIVU 1 Konteksti palautetaan oppilaiden mieliin käymällä Osan 1 johdanto uudelleen läpi. Kysymysten 1 ja 2 tarkoituksena on arvioida ovatko oppilaat ymmärtäneet
Vinkkejä opettajille ja odotetut tulokset SIVU 1 Konteksti palautetaan oppilaiden mieliin käymällä Osan 1 johdanto uudelleen läpi. Kysymysten 1 ja 2 tarkoituksena on arvioida ovatko oppilaat ymmärtäneet
Leena Keskitalo HYBRIDOOMASOLUN SISÄLTÄMÄN VASTA-AINEGEENIN MONISTAMINEN JA KLOONAAMINEN SEKVENSOINTIVEKTORIIN
 Leena Keskitalo HYBRIDOOMASOLUN SISÄLTÄMÄN VASTA-AINEGEENIN MONISTAMINEN JA KLOONAAMINEN SEKVENSOINTIVEKTORIIN HYBRIDOOMASOLUN SISÄLTÄMÄN VASTA-AINEGEENIN MONISTAMINEN JA KLOONAAMINEN SEKVENSOINTIVEKTORIIN
Leena Keskitalo HYBRIDOOMASOLUN SISÄLTÄMÄN VASTA-AINEGEENIN MONISTAMINEN JA KLOONAAMINEN SEKVENSOINTIVEKTORIIN HYBRIDOOMASOLUN SISÄLTÄMÄN VASTA-AINEGEENIN MONISTAMINEN JA KLOONAAMINEN SEKVENSOINTIVEKTORIIN
Eliömaailma. BI1 Elämä ja evoluutio Leena Kangas-Järviluoma
 Eliömaailma BI1 Elämä ja evoluutio Leena Kangas-Järviluoma Aitotumalliset l. eukaryootit Esitumalliset l. prokaryootit kasvit arkit alkueliöt sienet bakteerit eläimet Eliökunnan sukupuu Tumattomat eliöt
Eliömaailma BI1 Elämä ja evoluutio Leena Kangas-Järviluoma Aitotumalliset l. eukaryootit Esitumalliset l. prokaryootit kasvit arkit alkueliöt sienet bakteerit eläimet Eliökunnan sukupuu Tumattomat eliöt
Solun toiminta. II Solun toiminta. BI2 II Solun toiminta 7. Fotosynteesi tuottaa ravintoa eliökunnalle
 Solun toiminta II Solun toiminta 7. Fotosynteesi tuottaa ravintoa eliökunnalle 1. Avainsanat 2. Fotosynteesi eli yhteyttäminen 3. Viherhiukkanen eli kloroplasti 4. Fotosynteesin reaktiot 5. Mitä kasvit
Solun toiminta II Solun toiminta 7. Fotosynteesi tuottaa ravintoa eliökunnalle 1. Avainsanat 2. Fotosynteesi eli yhteyttäminen 3. Viherhiukkanen eli kloroplasti 4. Fotosynteesin reaktiot 5. Mitä kasvit
Vastaa lyhyesti selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1 1) Tunnista molekyylit (1 piste) ja täytä seuraava taulukko (2 pistettä) a) b) c) d) a) Syklinen AMP (camp) (0.25) b) Beta-karoteeni (0.25 p) c) Sakkaroosi (0.25 p) d) -D-Glukopyranoosi (0.25 p) 2 Taulukko.
1 1) Tunnista molekyylit (1 piste) ja täytä seuraava taulukko (2 pistettä) a) b) c) d) a) Syklinen AMP (camp) (0.25) b) Beta-karoteeni (0.25 p) c) Sakkaroosi (0.25 p) d) -D-Glukopyranoosi (0.25 p) 2 Taulukko.
Luennon 5 oppimistavoitteet. Soluseinän biosynteesi. Puu-19.210 Puun rakenne ja kemia. Solun organelleja. Elävä kasvisolu
 Luennon 5 oppimistavoitteet Soluseinän biosynteesi Puu-19.210 Puun rakenne ja kemia saat listata puuaineksen muodostumisen vaiheet. Ymmärrät, kuinka soluseinän tapahtuu. saat lyhyesti kuvata soluseinän
Luennon 5 oppimistavoitteet Soluseinän biosynteesi Puu-19.210 Puun rakenne ja kemia saat listata puuaineksen muodostumisen vaiheet. Ymmärrät, kuinka soluseinän tapahtuu. saat lyhyesti kuvata soluseinän
KOULUTUSOHJELMA Sukunimi: 18.5.2016 Etunimet: Nimikirjoitus: BIOLOGIA (45 p) Valintakoe klo 9.00-13.00
 Valintakoe klo 9.00-13.00") BIOLÄÄKETIETEEN Henkilötunnus: - KOULUTUSOHJELMA Sukunimi: 18.5.2016 Etunimet: Nimikirjoitus: BIOLOGIA (45 p) Valintakoe klo 9.00-13.00 Kirjoita selvästi nimesi ja muut henkilötietosi niille varattuun
BIOLÄÄKETIETEEN Henkilötunnus: - KOULUTUSOHJELMA Sukunimi: 18.5.2016 Etunimet: Nimikirjoitus: BIOLOGIA (45 p) Valintakoe klo 9.00-13.00 Kirjoita selvästi nimesi ja muut henkilötietosi niille varattuun
Muuttumaton genomi? Genomin ylläpito. Jakson luennot. Luennon sisältö DNA:N KAHDENTUMINEN ELI REPLIKAATIO
 Muuttumaton genomi? Genomin ylläpito SNP 14.1.2013 Tiina Immonen Biolääketieteen laitos Biokemia ja kehitysbiologia Jakson luennot Mitä on genomilääketiede? Dan Lindholm Genomin ylläpito Tiina Immonen
Muuttumaton genomi? Genomin ylläpito SNP 14.1.2013 Tiina Immonen Biolääketieteen laitos Biokemia ja kehitysbiologia Jakson luennot Mitä on genomilääketiede? Dan Lindholm Genomin ylläpito Tiina Immonen
Avainsanat: perimä dna rna 5`-ja 3`-päät replikaatio polymeraasientsyymi eksoni introni promoottori tehostajajakso silmukointi mutaatio
 Avainsanat: perimä dna rna 5`-ja 3`-päät replikaatio polymeraasientsyymi eksoni introni promoottori tehostajajakso silmukointi mutaatio Perinnöllinen informaatio sijaitsee dna:ssa eli deoksiribonukleiinihapossa
Avainsanat: perimä dna rna 5`-ja 3`-päät replikaatio polymeraasientsyymi eksoni introni promoottori tehostajajakso silmukointi mutaatio Perinnöllinen informaatio sijaitsee dna:ssa eli deoksiribonukleiinihapossa
DNA Tiina Immonen, FT, yo-lehtori HY Lääketieteellinen tiedekunta Biokemia ja kehitysbiologia
 DNA 18.4.2016 Tiina Immonen, FT, yo-lehtori HY Lääketieteellinen tiedekunta Biokemia ja kehitysbiologia Koordinaattori, Master s Degree Programme in Translational Medicine (TRANSMED) 1 Sisältö DNA:n rakenne
DNA 18.4.2016 Tiina Immonen, FT, yo-lehtori HY Lääketieteellinen tiedekunta Biokemia ja kehitysbiologia Koordinaattori, Master s Degree Programme in Translational Medicine (TRANSMED) 1 Sisältö DNA:n rakenne
Vasta-ainemääritys. Johdanto. www.edu.fi/biogeeni
 Vasta-ainemääritys Johdanto Vasta-ainemääritys (engl. immunoassay) perustuu spesifisen vasta-aineen (engl. antibody) sitoutumiseen mitattavaan antigeeniin (engl. antigen). Menetelmän etuja ovat suuri herkkyys
Vasta-ainemääritys Johdanto Vasta-ainemääritys (engl. immunoassay) perustuu spesifisen vasta-aineen (engl. antibody) sitoutumiseen mitattavaan antigeeniin (engl. antigen). Menetelmän etuja ovat suuri herkkyys
Väärin, Downin oireyhtymä johtuu ylimääräisestä kromosomista n.21 (trisomia) Geeni s. 93.
 Geeni s. 93.") 1 I) Ovatko väittämät oikein (O) vai väärin (V)? Jos väite on mielestäsi väärin, perustele se lyhyesti väittämän alla oleville riveille. O/V 1.2. Downin oireyhtymä johtuu pistemutaatista fenyylialaniinin
1 I) Ovatko väittämät oikein (O) vai väärin (V)? Jos väite on mielestäsi väärin, perustele se lyhyesti väittämän alla oleville riveille. O/V 1.2. Downin oireyhtymä johtuu pistemutaatista fenyylialaniinin
Agaroosigeelielektroforeesi (AGE)
") Agaroosigeelielektroforeesi (AGE) Johdanto Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNA/RNA -jaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Agaroosigeelielektroforeesin
Agaroosigeelielektroforeesi (AGE) Johdanto Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNA/RNA -jaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Agaroosigeelielektroforeesin
Genomi-ilmentyminen Genom expression (uttryckning) Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia
 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia") Genomi-ilmentyminen Genom expression (uttryckning) DNA RNA 7.12.2017 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia Osaamistavoitteet Lärandemål Luennon jälkeen ymmärrät pääperiaatteet
Genomi-ilmentyminen Genom expression (uttryckning) DNA RNA 7.12.2017 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia Osaamistavoitteet Lärandemål Luennon jälkeen ymmärrät pääperiaatteet
Biokemian menetelmät I kurssi, työselostukset, kevät 2016.
 Biokemian menetelmät I kurssi, työselostukset, kevät 2016. DEADLINET: työselostus tulostettuna paperille Työ 3: To 24.3.2016 klo 15:00 KE1132:n palautuspiste tai BMTK:n Työ 2: Pe 1.4.2016 klo 16:00 KE1132:n
Biokemian menetelmät I kurssi, työselostukset, kevät 2016. DEADLINET: työselostus tulostettuna paperille Työ 3: To 24.3.2016 klo 15:00 KE1132:n palautuspiste tai BMTK:n Työ 2: Pe 1.4.2016 klo 16:00 KE1132:n
Materiaalinäytteiden qpcr-tulosten tulkinnasta
 Materiaalinäytteiden qpcr-tulosten tulkinnasta Helena Rintala ja Teija Meklin Sisäilmastoseminaari 13.3.2014 Taustaa qpcr (kvantitatiivinen PCR) on nopea menetelmä mikrobien toteamiseen Käytetty paljon
Materiaalinäytteiden qpcr-tulosten tulkinnasta Helena Rintala ja Teija Meklin Sisäilmastoseminaari 13.3.2014 Taustaa qpcr (kvantitatiivinen PCR) on nopea menetelmä mikrobien toteamiseen Käytetty paljon
KEMIA. Kemia on tiede joka tutkii aineen koostumuksia, ominaisuuksia ja muuttumista.
 KEMIA Kemia on tiede joka tutkii aineen koostumuksia, ominaisuuksia ja muuttumista. Kemian työturvallisuudesta -Kemian tunneilla tutustutaan aineiden ominaisuuksiin Jotkin aineet syttyvät palamaan reagoidessaan
KEMIA Kemia on tiede joka tutkii aineen koostumuksia, ominaisuuksia ja muuttumista. Kemian työturvallisuudesta -Kemian tunneilla tutustutaan aineiden ominaisuuksiin Jotkin aineet syttyvät palamaan reagoidessaan
PIKORNAVIRUSGENOMIN MONISTUS RT-PCR- MENETELMÄLLÄ
 Opinnäytetyö (AMK) Bio- ja elintarviketekniikan koulutusohjelma Biotekniikka 2018 Tapio Metz PIKORNAVIRUSGENOMIN MONISTUS RT-PCR- MENETELMÄLLÄ OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio-
Opinnäytetyö (AMK) Bio- ja elintarviketekniikan koulutusohjelma Biotekniikka 2018 Tapio Metz PIKORNAVIRUSGENOMIN MONISTUS RT-PCR- MENETELMÄLLÄ OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio-
PCR:n laadunvarmistus. Saija Hallanvuo Elintarvike- ja rehumikrobiologian tutkimusyksikkö/ Elintarvikemikrobiologiajaosto
 PCR:n laadunvarmistus Saija Hallanvuo Elintarvike- ja rehumikrobiologian tutkimusyksikkö/ Elintarvikemikrobiologiajaosto Ajankohtaista laboratoriorintamalla 11.10.2012 1 Laadunvarmistus Validointi Henkilökunnan
PCR:n laadunvarmistus Saija Hallanvuo Elintarvike- ja rehumikrobiologian tutkimusyksikkö/ Elintarvikemikrobiologiajaosto Ajankohtaista laboratoriorintamalla 11.10.2012 1 Laadunvarmistus Validointi Henkilökunnan
Yoshinori Ohsumille Syntymäpaikka Fukuoka, Japani 2009 Professori, Tokyo Institute of Technology
 Lääketieteen Nobel-palkinto 2016 Yoshinori Ohsumille hänen autofagian mekanismeja koskevista löydöistään. Yoshinori Ohsumi 1945 Syntymäpaikka Fukuoka, Japani 2009 Professori, Tokyo Institute of Technology
Lääketieteen Nobel-palkinto 2016 Yoshinori Ohsumille hänen autofagian mekanismeja koskevista löydöistään. Yoshinori Ohsumi 1945 Syntymäpaikka Fukuoka, Japani 2009 Professori, Tokyo Institute of Technology
Solun perusrakenne I Solun perusrakenne. BI2 I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja
 Solun perusrakenne I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja 1. Avainsanat 2. Solut tuottavat entsyymejä katalyyteiksi 3. Entsyymien rakenne ja toiminta 4. Entsyymit vaativat toimiakseen
Solun perusrakenne I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja 1. Avainsanat 2. Solut tuottavat entsyymejä katalyyteiksi 3. Entsyymien rakenne ja toiminta 4. Entsyymit vaativat toimiakseen
Päästä varpaisiin. Tehtävät. Ratkaisut. Päivitetty 8.4.2013 ISBN 978-951-37-6416-6, 978-951-37-6417-3, 978-951-6418-0. Sisällys (ratkaisut) Johdanto
 Johdanto") OPETTAJAN AINEISTO Käyttöehdot Päästä varpaisiin Ihmisen anatomia ja fysiologia Eliisa Karhumäki Mari Kärkkäinen (os. Lehtonen) Päivitetty 8.4.2013 ISBN 978-951-37-6416-6, 978-951-37-6417-3, 978-951-6418-0
OPETTAJAN AINEISTO Käyttöehdot Päästä varpaisiin Ihmisen anatomia ja fysiologia Eliisa Karhumäki Mari Kärkkäinen (os. Lehtonen) Päivitetty 8.4.2013 ISBN 978-951-37-6416-6, 978-951-37-6417-3, 978-951-6418-0
TÄS ON PROTSKUU! Missä yhteyksissä olet törmännyt sanaan proteiini tai valkuaisaine?
 TÄS ON PROTSKUU! KOHDERYHMÄ: Työ soveltuu parhaiten yläkouluun kurssille elollinen luonto ja yhteiskunta, sekä lukioon kurssille KE1. KESTO: Työ koostuu kahdesta osasta: n. 30 min/osa. MOTIVAATIO: Mitä
TÄS ON PROTSKUU! KOHDERYHMÄ: Työ soveltuu parhaiten yläkouluun kurssille elollinen luonto ja yhteiskunta, sekä lukioon kurssille KE1. KESTO: Työ koostuu kahdesta osasta: n. 30 min/osa. MOTIVAATIO: Mitä
Säteilyvaikutuksen synty. Erikoistuvien lääkärien päivät 25 26.1.2013 Kuopio
 Säteilyvaikutuksen synty Erikoistuvien lääkärien päivät 25 26.1.2013 Kuopio Säteilyn ja biologisen materian vuorovaikutus Koska ihmisestä 70% on vettä, todennäköisin (ja tärkein) säteilyn ja biologisen
Säteilyvaikutuksen synty Erikoistuvien lääkärien päivät 25 26.1.2013 Kuopio Säteilyn ja biologisen materian vuorovaikutus Koska ihmisestä 70% on vettä, todennäköisin (ja tärkein) säteilyn ja biologisen
Ohjeita opettamiseen ja odotettavissa olevat tulokset SIVU 1
 Ohjeita opettamiseen ja odotettavissa olevat tulokset SIVU 1 Toiminta aloitetaan johdattelulla. Tarkoituksena on rakentaa konteksti oppilaiden tutkimukselle ja kysymykselle (Boldattuna oppilaiden työohjeessa),
Ohjeita opettamiseen ja odotettavissa olevat tulokset SIVU 1 Toiminta aloitetaan johdattelulla. Tarkoituksena on rakentaa konteksti oppilaiden tutkimukselle ja kysymykselle (Boldattuna oppilaiden työohjeessa),
Tuija Solismaa IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS
 Tuija Solismaa IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS RAPORTIN NIMIÖSIVU IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS
Tuija Solismaa IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS RAPORTIN NIMIÖSIVU IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS