Miten fylogenioita voidaan käyttää hyväksi eliökunnan historian tutkimisessa?
|
|
|
- Ari-Pekka Halttunen
- 7 vuotta sitten
- Katselukertoja:
Transkriptio
1 Miten fylogenioita voidaan käyttää hyväksi eliökunnan historian tutkimisessa? -fylogeneettisen puun käsite -fylogenioiden tavalliset rakennusmenetelmät (parsimonia, UPGMA, neighbor joining, maximum likelihood) -puun luotettavuuden arviointi bootstrap-menetelmällä -mitä sekvenssejä ja mitä rakennusmenetelmää käytetään?

2 4. Molekyylifylogeniat 4.1. Mihin molekyylifylogenioita käytetään? 4.2. Molekyylidatan edut 4.3. Terminologiaa 4.4. Fylogeniamenetelmät Parsimoniamenetelmä Etäisyysmatriisimenetelmät -UPGMA -Neighbor-joining Maximum likelihood Juuri 4.5. Fylogenian luotettavuuden arviointi 4.6. Geenipuu ja lajipuu 4.7. Kladistit ja fenetistit 4.8. Yhteenveto menetelmistä
3 4.1. Mihin molekyylifylogenioita käytetään? Fylogeneettisen analyysin tavoitteena on 1) selvittää lajien (tai muiden taksonien) historialliset suhteet 2) yksilöiden tai yleensä yksittäisten DNA-sekvenssien historiallisten suhteiden selvittäminen 3) selvittää duplikoituneiden sekvenssien ja geeniperheen jäsenten väliset historialliset suhteet

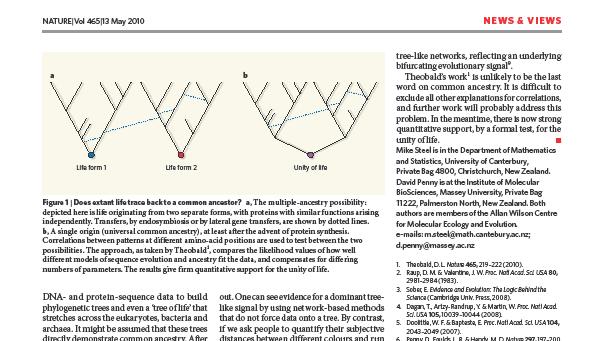
4 Tree of life Elämän puu
5 Päälinjojen suhteista on vaihtoehtoisia hypoteeseja joista keskustellaan: The "archaea tree": The "eocyte tree":

6
7 HIV-isolaattien fylogenia (nukleotidisekvenssidata)

8 Nature Apr 22;428(6985):820. Origin of AIDS: contaminated polio vaccine theory refuted. Worobey M, Santiago ML, Keele BF, Ndjango JB, Joy JB, Labama BL, Dhed'A BD, Rambaut A, Sharp PM, Shaw GM, Hahn BH.
9 87 taskurottayksilön mitokondriosekvenssin fylogenia Avise ym. 1979
10 Ihmisen globiinigeeniperheen fylogenia GLs280

11 MADS-box-geenien fylogenia Pun.=eläimet ja sienet Musta=Arabidopsis Haarojen pituudet suhteessa a-a muutoksiin Suurin osa kasveilla tyyppiä II K-domaini vain kasvilinjassa Medard Ng and Yanofsky 2001
12 Kotitehtävä 1 (harj. 3) a) Etsi kirjastosta jostain tieteellisestä artikkelista (Molecular Phylogeny and Evolution, Molecular Biology and Evolution, Heredity, Genetics) fylogeneettinen puu. Katso 1)miksi se on tehty (mihin tieteelliseen kysymykseen se on vastaus) 2)millä menetelmällä se on tehty Geeni/geeniperhe DNA/RNA/aa tuma/mt/? Laji/lajiryhmä substituutiomalli puunrakennusmenete lmä
13 Kotitehtävä 2 (harj. 3) c) Käy tutkimassa seuraavat puiden käyttöesimerkit: Using trees to make predictions about fossils: The whale's ankle Using trees to learn about the evolution of complex features: The striped cichlid Using trees to make predictions about poorly-studied species: A new drug Using trees to learn about the order of evolution: The spider's web Using trees to learn about the evolution of diversity: The beetles' diet -parsimoniaan perustuvaa päättely
14 4.2. Molekyylidatan edut Molekyylidata: -aminohapposekvenssit ( luvut) -allotsyymit -DNA-DNA-hybridisaatio -immunologiset menetelmät -DNA-sekvenssit 1970-l. lopulta lähtien -genomisekvenssit
15 John Ray, 1962 (Bromham 2008)
16 Molekyylidata: -100% periytyvää - on-off muuntelu (ei-jatkuva) - homologian määrittäminen (=sekv. linjaus) helppoa -laskennalliset menetelmät edistyneitä -sopii kaukaisillekin sukulaisille -paljon ominaisuuksia Morfologiset ja fysiologiset ominaisuudet: -vain osin periytyvää -jatkuva muuntelu -homologisten ominaisuuksien määrittäminen on vaikeaa -laskennalliset menetelmät eivät edistyneitä -eisovikaukaisillesukulaisille -vähän ominaisuuksia -kun DNA näytettä ei saa (historialliset taksonit)
17 4.3. Terminologiaa -vain haarautumisjärjestys (topologia): haarojen pituudet eivät kuvaa mitään A B C D -haarojen pituudet suhteessa A B A B C D C D kuluneeseen aikaan tapahtuneisiin muutoksiin
18 Bromham 2008
19 -a, b, c, d operationaalisia taksonomisia yksiköitä (OTU) -f, g, h hypoteettisia taksonomisia yksiköitä; Sisäinen haara (internal branch) g h Solmu (node) f h g f a b c d a b c d Ulkoinen haara (external branch)
20 Solmusta voi lähteä kaksi haaraa (dikotomia) (bifurcating) tai monta haaraa (polytomia) (multifurcating); -monta lajiutumista tapahtui samaan aikaan/ menetelmä ei erota haarautumisjärjestystä (esim. liian vähän dataa).
21 Puu voi olla juureton tai juurrettu Juurellinen: evoluutiolla suunta a a b c d Juureton: evoluutiolla ei suuntaa a b c d a b c d c d a b a b d c c d b a
22 Mahdollisten puiden määrä: Sekvenssejä: Juurettomia puita: Juurellisia puita: Usein riittää pelkkä juureton puu, esim. kun selvitetään monofyleettisiä ryhmiä.
23 Mono- ja parafyleettiset ryhmät Monofyleettinen ryhmä: kaikki taksonit jotka polveutuvat samasta esiisästä (+ esi-isä). Ryhmällä on yhteinen esi-isä, joka ei ole esi-isä ryhmän ulkopuolisille jäsenille. Esim. krokotiilit ja linnut Parafyleettinen ryhmä: yhteinen esi-isä on myös jonkun toisen taksonomisen ryhmän esi-isä. Esim. matelijat L i n n u t LINNUT KROKOTIILIT KÄÄRMEET JA SISILISKOT MATELIJAT KILPIKONNAT NISÄKKÄÄT
24 Newickin formaatti: tapa puun rakenteen kuvaamiseen. Kukin sulkumerkkipari sisältää monofyleettisen ryhmän A F B C ((((A,B)C)D)E) A E D E F B D 2,1 C A ((A,B),(C,D),(E,F)) 2,5 1,2 E 1,1 1,0 1,0 ((A:2.5,B:1.1):1.0,(C:0.8,D:2.0):1.1,(E:1.2,F:2.1):1.0) B 1,1 2,0 D C 0,8
25 4.4. Fylogeniamenetelmät 1) Parsimoniamenetelmä Yhteiset mutaatiot kertovat yhteisestä alkuperästä > perustuu ominaisuuksien tiloihin (character state) (linjatun DNA:n nukleotipaikka (tilat: A, T, C, G), prot. aminohappopaikka (tilat: 20 aminohappoa), transposoni (tilat: on - ei ole), insertio/deleetio (indel) DNA tai aminohapposekvenssissä (tilat: on ei ole) 2) Etäisyysmatriisimenetelmät Samankaltaisuus kertoo yhteisestä alkuperästä > menetelmät perustuvat parittaisiin etäisyysmittoihin (esim. DNAsekvensseissä korvautumisten määrä K parittaisten sekvenssien välillä) 3) Maximum likelihood (ML) - tyyppiset menetelmät; voivat käyttää sekä ominaisuuksien tiloja että parittaisia etäisyyksiä; maksimoivat datan todennäköisyyden evolutiivista mallia käyttäen.
26 4.4.1 Parsimoniamenetelmä Parsimoniaperiaate: -minimoidaan tapahtuneiden muutosten määrä ->parsimonisin puu: selittyy pienimmällä määrällä mutaatioita Tilojen muutoksessa voi olla suunta; esim. DNA:n nukleotidivaihdoksissa ei ole suuntaa Tilojen välillä voi olla tietty määrä muutoksia; esim. tietyn aminohapon muutos toiseksi voi vaatia useita muutoksia.
27 Parsimoniamenetelmässä yhteiset mutaatiot kertovat yhteisestä alkuperästä. Se perustuu fylogeneettisesti informatiivisiin muutoksiin = muutoksiin jotka jakavat aineiston kahteen ryhmään. Oranki atgcaagtgcagccatcgacgacttacgacgaccgccagt Ihminen.t.t.agc...a.g.cc...c.cgca.t.gt...g.. Gorilla.c.t.ggc...a.g.tt...a.tgcg.t.at...a.. Simpans.t.c.act...a.g.cc...a.cgta.t.at...g.. Fylogeneettisesti informatiiviset muutokset Yksittäismutaatiot (singletons)
28 Oranki atgcaagtgcagccatcgacgacttacgacgaccgccagt Ihminen.t.t.agc...a.g.cc...c.cgca.t.gt...g.. Gorilla.c.t.ggc...a.g.tt...a.tgcg.t.at...a.. Simpans.t.c.act...a.g.cc...a.cgta.t.at...g Mahdolliset juurettomat topologiat: I G I S I S S O G O O G Paikkojen lkm jotka tukevat kutakin ryhmittelyä
29 4. (8.) I S G O I S G O I S G O C T T C t t C T T C t c C T C T c c I S G O I S G O I S G O T C T C c t T C T C c c T C C T c c I S G O I S G O I S G O C C A A c c C C A A c c C C A A c a 16.(24.,25.,26.) 23.
30 Summataan muutokset yli kaikkien informatiivisten paikkojen kullekin mahdolliselle puulle erikseen: I G I S I S S O G O O G 2 paikkaa x 2 muutosta + 4 paikkaa x 1 muutos +1 paikka x 2 muutosta = 10 2 paikkaa x 1 muutosta + 4 paikkaa x 2 muutos +1 paikka x 2 muutosta = 12 2 paikkaa x 2 muutosta + 4 paikkaa x 2 muutos +1 paikka x 1 muutosta = 13 Paras puu = parsimonisin puu: selittyy pienimmällä määrällä mutaatioita Kun OTUja on yli neljä, parsimonisin puu ei ole välttämättä se jota tukee suurin määrä informatiivisia paikkoja, koska kukin informatiivinen paikka ei välttämättä enää tue vain yhtä topologiaa.
31 Parsimoniamenetelmä tuottaa pelkän topologian ilman juurta ja haarojen pituuksia. L i n n u t Parsimoniapuun ongelmana on homoplasia kun muutoksia on paljon. Eri nukleotidipaikkoja tai muutoksia on mahdollista painottaa eri tavoin; esim. jos on odotettavissa paljon homoplasioita, näitä painotetaan vähemmän. -esim. 1., 2. vs. 3. paikka kodonissa -esim. transversiot vs. transitiot
32 > homoplasia
33 ATG GCT ACC ATG GCC GCC ATG GCT ACT ATA GCC GCC ATG GCC GCC ATA GCT ACT ATG GCC ACT Symplesiomorfia: identtisyys perustuu siihen, että kumpikaan ei ole muuttunut. Synapomorfia: identtisyys joka seurausta yhdestä mutaatiosta. Homoplasia: identtisyys johtuu saman nukleotidin moninkertaisesta muuttumisesta (kaksi erillistä mutaatiota jotka tuottavat saman lopputuloksen)



34 Subst kpl Subst. osuus Hav. kpl Hav. osuus Od. osuus 1. paikka transitiot 2. paikka transver siot 3. paikka
35 Parsimoniamenetelmällä ei aina löydä parsimonisinta puuta Parsimoniamenetelmässä verrataan kaikkia mahdollisia juurettomia puita (exhaustive search): laskennallisesti raskas. Kun kaikkia mahdollisia puita ei voi verrata, käytetään oikopolkuja: Kun OTUja on > 12 branch and bound-menetelmä: lisätään puuhun taksoneita yksi kerrallaan (puun kokonaispituus kasvaa): verrataan kullakin askelella esim. Neighbor-joining menetelmällä saadun puun kokonaispituuteen (mutaatioiden kokon. määrään). Hylätään tämä polku jos pituus suurempi. Kun OTUja > 20 heuristic search: otetaan käsittelyyn vain sellaisia puita jotka muistuttavat topologialtaan neighbor-joining puuta. Muita: branch swapping: tutkitaan muunnelmia peruspuusta vaihtamalla läheisten haarojen päissä olevia taksoneita.
36 Branch and bound GL196
37 4.4.2 Etäisyysmatriisimenetelmät Jotkut menetelmät kuten DNA-DNA-hybridisaatio tai immunologiset menetelmät tuottavat suoraan etäisyysdataa. Monet menetelmät tuottavat dataa jossa kyseessä on ominaisuuden tilat (kuten DNA-sekvensointi) mutta datan voi muuttaa etäisyysdataksi. Etäisyysdataa ei voi muuttaa ominaisuusdataksi. DNA-sekvensseistä tehdään parittaisten etäisyyksien (K) matriisi käyttäen esim. Jukes-Cantorin, Kimuran 2 parametrin tai muuta moninkertaisten substituutioiden korjausta. Puunrakennusalgoritmeja on mm. UPGMA, Neigbor-joining
38 Oranki Ihminen Gorilla Simpanssi Paviaani atgcaagtgcagccatcgacgacttacgacgaccgccagt.t.t.agc...a.g.cc...c.cgca.t.gt...g...c.t.ggc...a.g.tt...a.tgcg.t.at...a...t.c.act...a.g.cc...a.cgta.t.at...g...tac.ggc.t.t.c.tg.g..ga.tgcg.caat..t.a.. Nukleotidierot p Jukes-Cantor etäisyys K I S G O P S G O P I ,167 0,268 0,383 0,627 S ,304 0,383 0,687 G ,343 0,383 O ,426 P -
39 UPGMA Unweighted pair-group method with arithmetic means Ryhmitetään sekvenssit joilla on pienin etäisyys Lasketaan kaikkien muiden keskietäisyys muodostuneeseen ryhmään ja jatketaan siitä Oletuksena on tasainen evoluutionopeus linjoissa ->Tuottaa puun jossa haarojen pituus on suhteessa aikaan ->Tuottaa juuren
40 S G O P I 0,167 0,268 0,383 0,627 S 0,304 0,383 0,687 G 0,343 0,383 O 0,426 G O P I+S 0,286 0,383 0,657 G 0,343 0,383 O 0,426 O P I+S+G 0,370 0,566 O 0,426 P I+S+G+O 0,531
41 Ihminen Simpanssi Gorilla Oranki Paviaani 0,25 0,20 0,15 0,10 0,05 0
42 Neighbor joining Etsitään vaiheittain naapurit siten että puun kokonaispituus (=kaikkien haarojen summa) minimoituu y x 3 x y x lasketaan puun kok. pituus kaikille vaihtoehtoisille puille jossa solmussa X on yhdistyneinä kaikki mahdolliset OTUt
43 Kun on löydetty lyhin puu, muodostetaan X:n yhdistämistä kahdesta OTUsta yhdistelmä-otu ja lasketaan muiden OTUjen ja yhdistelmä-otun väliset etäisyydet keskiarvona (kuten UPGMAssa). Etsitään taas lyhin puu mahdollisten topologioiden joukosta.
44 Neigbor joining puu edellisestä aineistosta Ihminen Simpanssi Gorilla Oranki Paviaani Menetelmä ei oleta että evoluutionopeus on sama kaikissa linjoissa -> Haarojen pituus on suhteessa tapahtuneisiin muutoksiin. -> ei saada juurta.
45 4.4.3 Suurimman todennäköisyyden (maksimum likelihood) menetelmät Likelihood = todennäköisyys havaita data (sekvenssit) olettaen tietty haarautumisjärjestys ja nukleotidisubstituutiomalli (esim. kahden parametrin malli) L=P(data puu) Lasketaan datan todennäköisyys kaikille mahdollisille puille, ja valitaan puu jonka todennäköisyys on suurin. Samalla arvioidaan puun haarojen pituudet. Vaatii tietokoneelta paljon. Bayesiläiset puunrakennusmenetelmät ovat samantapaisia, mutta likelihood:n asemesta todennäköisyys saadaan jakaumana.
46 GL199
47 4.4.3 Juuri -Useimmat menetelmät eivät tuota juurta -UPGMA tuottaa juuren, koska menetelmässä oletetaan tasainen evoluutionopeus) -Juuren saamiseksi fylogeniaan otetaan mukaan sopivan kaukainen taksoni, jonka tiedetään eronneen muista selvästi aiemmin esim. paleontologisten tietojen perusteella. Juuri laitetaan haaraan joka yhdistää ulkoryhmän (outgroup) muihin taksoneihin. Ulkoryhmän on hyvä olla riittävän erilainen mutta ei liian erilainen (ettei homoplasioiden takia saada esim. epäluotettavia etäisyyksiä tai väärää topologiaa). ulkoryhmä
48 Toinen menetelmä on asettaa juuri sen haaran puoliväliin joka erottaa kahden toisistaan kaukaisimman taksonin puoliväliin. Päätelty juuri
49 4.5. Fylogenian luotettavuuden arviointi Bootstrap-menetelmä: Tavoitteena on selvittää fylogeneettisellä menetelmällä päätellyn puun herkkyys pienille muutoksille havaintoaineistossa Otetaan sekvensseistä uusintaotos poimimalla satunnaisesti nukleotidipaikkoja takaisinpanolla Satunnaisotannan tuloksena saaduista sekvensseistä laaditaan uusi fylogenia samalla menetelmällä Uusintaotoksia ja fylogenioita tehdään esim kpl
50 Näyte Bootstrap-näytteet GAGGG AGGAC CCGAT CAAAA 2 GCGTG GGGAA CCGGA GAAAA 3 CAGAG AGAAA CAGAG TAAAC 4 CAAAG AGCAA CGAGT TAAAC 5 GCGGA CAGAA AAGAT TAAAT 1 GGGGAAAAGGCGGGGTCAAA 2 GGGGCGGGGGAGGGGAGAAA 3 CCCCAAAAAAAGGGGGTAAA 4 CCCCAAAAAAAGGGGGTAAA 5 GGGGCCCCGGAGGGGTTAAA Päätelty puu 2 1 AAAAGGGGACCCCAAAAAAA 2 CCCCGGGGAACCCGGAAAAA 3 AAAAGGAAAACAAAAAAACC 4 AAAAGGAACAACCAAAAACC 5 CCCCAAGGAAAAAAAAAATT... 1 GGGGGAGGAACCCCCCCAAA 2 GGGGGGGGAACCCCCCCAAA 3 GGGGGAGGAACCCCCAAAAA 4 AAAGGAGGCCAAAAACCAAA 5 GGGAACAAAAAAAAAAAAAA
51 Bootstrap-arvo: 3 1 Alkuperäisen päätellyssä sukupuussa esiintyvän ryhmän luotettavuutta kuvastaa se kuinka usein sama ryhmä esiintyy satunnaistetuissa uusissa puissa
52 Physarum Zea mays Euglena gracilis Trypanosoma brucei Naegleria gruberi Chlamydomonas reinhardtii Human Konsensus -puu (50%) Physarum Zea mays Euglena gracilis Trypanosoma brucei Naegleria gruberi Chlamydomonas reinhardtii Human
53 Physarum Zea mays Euglena gracilis Trypanosoma brucei Naegleria gruberi Chlamydomonas reinhardtii Human Konsensus -puu (90%) Physarum Zea mays Euglena gracilis Trypanosoma brucei Naegleria gruberi Chlamydomonas reinhardtii Human
54 4.6. Geenipuu ja lajipuu Lajin sisällä on polymorfiaa Geenin sekvenssien eroamisajankohta on sama tai kaukaisempi kuin lajien GL174
55 GL 175 -Geenipuun topologia voi olla erilainen kuin lajipuun -Kun kaksi perättäistä lajiutumista tapahtuu lyhyen ajan sisällä, väärän topologian saamiseen on suuri mahdollisuus. -Kun halutaan lajin fylogenia, luotettavimman tuloksen saa tutkimalla monta geeniä
56 4.7. Kladistit ja fenetistit Kladistit: kiinnostuneet evoluution kulusta; taksonien polveutumisjärjestyksestä eli kladogrammista (= fylogeneettisen juurellisen puun topologiasta). Parsimonia on tyypillinen kladistinen menetelmä. Fenetistit: kiinnostuneet olemassaolevien taksonien samankaltaisuuksista. Samankaltaisuudet ilmaistaan fenogrammilla. UPGMA tyypillinen feneettinen menetelmä.
57 4.8. Yhteenveto menetelmistä: Valitse geeni jonka evoluutionopeus on sopiva sille ryhmälle josta fylogeneettinen puu tehdään >homoplasioiden välttäminen >riittävät erot taksonien välillä Hitaasti kehittyvä alue kun on kyseessä kaukaiset taksonit: esim. Paljassiemeniset ja koppisiemenisten eroaminen, eubakteerien, eukaryoottien ja arkkien eroaminen. Nopeasti kehittyvä alue kun on kyseessä läheiset taksonit: esim. Brassicaceae-heimon lajien fylogenia, kädellisten fylogenia.
58 rrna mt Pieni suuri Käytettävän geenin, geenialueen tai muutoksen evoluutionopeus Avise 1994
59 Valitse paikat tai muutokset joissa on sopiva korvautumisnopeus: transitiot vs. transversiot kodonin ja 3. paikka Otetaanko aminohappoa muuttavat muutokset mukaan? synonyymiset vs. ei- synonyymiset muutokset ei-degeneroituneet vs. 4-kertaisesti degeneroituneet paikat -Huomioi myös emäskoostumuksen erot taksonien välillä.
60 Esim. tuman ja organellien geenien evoluutionopeuksien eroja eläimillä ja kasveilla Eläinten mitokondrio -n bp -DNA-sekvenssin syn. evoluutionopeus (5.7 x 10-8/v.) on 10- kertainen verrattuna tuman DNA:han. -> läheisille taksoneille -geenijärjestys ja lkm on vakaa -geenit ovat intronittomia -ihmismitokondrion kontrollialueessa paljon transitioita Transitiot : transversiot 15,7 : 1
61 Kasvien mitokondrioiden -koko vaihtelee paljon ( bp) lajista toiseen -geenisisältö vakaa -uudelleenjärjestelyjä on paljon -sekvenssievoluutio on erittäin hidasta verrattuna tumaan. kloroplastit -suuria ( bp) -geenisisältö vaihtelee paljon lajista toiseen -sekvenssievoluutio on hidasta verrattuna tumaan Synonyymisiä substituutioita (Ks) maissin ja vehnä/ohran välillä: Tuma 0, Kloroplasti 0,17 + 0,01 Mitokondrio 0,03 + 0,01
62 Esim. Havaitset seuraavat erilaistumisasteet puulajien välillä (Jukes-Cantor etäisyyksiä) P. sylvestris ja P. taeda 0.03, P. sylvestris ja Picea abies 0.14, P. taeda ja P. abies Pseudotsuga menziesii ja Picea abies 0.20 P. menziesii ja P. sylvestris 0.25 P. taeda ja P. menziesii Piirrä fylogeneettinen puu (UPGMA) näistä lajeista. P. menziesii tiedetään ulkoryhmäksi. Onko evoluutionopeus kuusen Picea ja männyn Pinus linjoissa erilainen? Jos oletat tätä tehtävän osaa varten, että evoluutionopeus on ollut sama kummassakin (kuusen ja männyn) linjassa, milloin arvioit kahden mäntylajin eronneen toisistaan? Oleta että männyn (Pinus) ja kuusen (Picea) suvut ovat eronneet noin 140 miljoonaa vuotta sitten.
13.11. Tulosten arviointi. tulosten arviointi. voimmeko luottaa saamiimme tuloksiin?
 13.11. tulosten arviointi Tulosten arviointi voimmeko luottaa saamiimme tuloksiin? onko osa saaduista tuloksista sattumanvaraisia? mitkä OSAT puusta ovat luotettavimpia? 1 KONSENSUSDIAGRAMMI Useita yhtä
13.11. tulosten arviointi Tulosten arviointi voimmeko luottaa saamiimme tuloksiin? onko osa saaduista tuloksista sattumanvaraisia? mitkä OSAT puusta ovat luotettavimpia? 1 KONSENSUSDIAGRAMMI Useita yhtä
Evoluutiovoimat. Mikä on mutaation, valinnan ja sattuman merkitys evoluutiossa?
 Evoluutiovoimat Mikä on mutaation, valinnan ja sattuman merkitys evoluutiossa? -sattuman sysäily: populaatiokoon vaikutus -valinta: positiivinen, tasapainottava ja negatiivinen -mutaatiot: neutraalien,
Evoluutiovoimat Mikä on mutaation, valinnan ja sattuman merkitys evoluutiossa? -sattuman sysäily: populaatiokoon vaikutus -valinta: positiivinen, tasapainottava ja negatiivinen -mutaatiot: neutraalien,
Molekyylievoluutio. (753327A, S), 4 op syksy Photo Marie-Hélène Müller
, 4 op syksy Photo Marie-Hélène Müller") Molekyylievoluutio (753327A, 753627S), 4 op syksy 2011 Photo Marie-Hélène Müller Molekyylievoluutio (753327A, 753627S), 4 op, syksy 2011 -Helmi Kuittinen p. 553 1803, PT312, helmi.kuittinen@oulu.fi harj.
Molekyylievoluutio (753327A, 753627S), 4 op syksy 2011 Photo Marie-Hélène Müller Molekyylievoluutio (753327A, 753627S), 4 op, syksy 2011 -Helmi Kuittinen p. 553 1803, PT312, helmi.kuittinen@oulu.fi harj.
Sekvenssievoluutio ja fylogeniat
 Sekvenssievoluutio ja fylogeniat Miksi ja miten nukleotidikorvautumiset lasketaan havaittujen sekvenssierojen perusteella? -sekvenssien linjaus -mitä eroa on eroilla ja korvautumisilla - nukleotidikorvautumisten
Sekvenssievoluutio ja fylogeniat Miksi ja miten nukleotidikorvautumiset lasketaan havaittujen sekvenssierojen perusteella? -sekvenssien linjaus -mitä eroa on eroilla ja korvautumisilla - nukleotidikorvautumisten
Fylogeneettiset puut. Fylogeneettiset puut. UPGMA: esimerkki 2/2 UPGMA
 ylogeneettiset puut ylogeneettisen puun rakentaminen koostuu seuraavista vaiheista ) atan valinta (sekvenssi,piirredata) ) Sekvenssien linjaus 3) Puun rakentamismenetelmän/menetelmien valinta: - etäisyysmenetelmät
ylogeneettiset puut ylogeneettisen puun rakentaminen koostuu seuraavista vaiheista ) atan valinta (sekvenssi,piirredata) ) Sekvenssien linjaus 3) Puun rakentamismenetelmän/menetelmien valinta: - etäisyysmenetelmät
Molekyylisystematiikka 1.osa
 Molekyylisystematiikka 1.osa Johdanto Käsitteet Sukulaisuuksien esittäminen eri formaateissa Puut: eri tavat muodostaa puu, algoritmeja, ohjelmistoja, esimerkki Petri Törönen Vanha materiaali: Päivi Onkamo,
Molekyylisystematiikka 1.osa Johdanto Käsitteet Sukulaisuuksien esittäminen eri formaateissa Puut: eri tavat muodostaa puu, algoritmeja, ohjelmistoja, esimerkki Petri Törönen Vanha materiaali: Päivi Onkamo,
Sekvenssievoluutio ja fylogeniat
 Sekvenssievoluutio ja fylogeniat Miksi sekvenssien evoluutionopeus vaihtelee? -erot korvautumisnopeudessa geenien tai geenin eri osien välillä, -erot korvautumisnopeudessa eri fylogeneettisten linjojen
Sekvenssievoluutio ja fylogeniat Miksi sekvenssien evoluutionopeus vaihtelee? -erot korvautumisnopeudessa geenien tai geenin eri osien välillä, -erot korvautumisnopeudessa eri fylogeneettisten linjojen
Molekyylisystematiikka, 2. osa
 Molekyylisystematiikka, 2. osa Parsimoniamenetelmä, hyvät ja huonot puolet Plussat: Helppo ymmärtää, ei oleta mitään tiettyä evolutiivista mallia Voidaan osoittaa että löytää varmaasti parhaan puun Hypotetisoi
Molekyylisystematiikka, 2. osa Parsimoniamenetelmä, hyvät ja huonot puolet Plussat: Helppo ymmärtää, ei oleta mitään tiettyä evolutiivista mallia Voidaan osoittaa että löytää varmaasti parhaan puun Hypotetisoi
6.4. Genomin koon evoluutio Genomin koko vaihtelee
 6.4. Genomin koon evoluutio 6.4.1. Genomin koko vaihtelee C-arvo: genomin haploidi koko pg:na 1 pg = 0.98 x 10 9 bp = 1 milj. kb = 1000 Mb (ero: geneettinen genomin koko (cm)) Missäkohtaa genomiaon kokoeroja?
6.4. Genomin koon evoluutio 6.4.1. Genomin koko vaihtelee C-arvo: genomin haploidi koko pg:na 1 pg = 0.98 x 10 9 bp = 1 milj. kb = 1000 Mb (ero: geneettinen genomin koko (cm)) Missäkohtaa genomiaon kokoeroja?
Evoluutiopuu. Aluksi. Avainsanat: biomatematiikka, päättely, kombinatoriikka, verkot. Luokkataso: 6.-9. luokka, lukio
 Evoluutiopuu Avainsanat: biomatematiikka, päättely, kombinatoriikka, verkot Luokkataso: 6.-9. luokka, lukio Välineet: loogiset palat, paperia, kyniä Kuvaus: Tehtävässä tutkitaan bakteerien evoluutiota.
Evoluutiopuu Avainsanat: biomatematiikka, päättely, kombinatoriikka, verkot Luokkataso: 6.-9. luokka, lukio Välineet: loogiset palat, paperia, kyniä Kuvaus: Tehtävässä tutkitaan bakteerien evoluutiota.
Evoluutiovoimat. Ydinkysymykset. Mikä on mutaation, valinnan ja sattuman merkitys evoluutiossa?
 Evoluutiovoimat Ydinkysymykset Mikä on mutaation, valinnan ja sattuman merkitys evoluutiossa? -sattuman sysäily: populaatiokoon vaikutus -valinta: positiivinen, tasapainottava ja negatiivinen -mutaatiot:
Evoluutiovoimat Ydinkysymykset Mikä on mutaation, valinnan ja sattuman merkitys evoluutiossa? -sattuman sysäily: populaatiokoon vaikutus -valinta: positiivinen, tasapainottava ja negatiivinen -mutaatiot:
DNA RNA proteiinit transkriptio prosessointi translaatio regulaatio
 replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi translaatio
replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi translaatio
S Laskennallinen systeemibiologia
 S-114.2510 Laskennallinen systeemibiologia 3. Harjoitus 1. Koska tilanne on Hardy-Weinbergin tasapainossa luonnonvalintaa lukuunottamatta, saadaan alleeleista muodostuvien eri tsygoottien genotyyppifrekvenssit
S-114.2510 Laskennallinen systeemibiologia 3. Harjoitus 1. Koska tilanne on Hardy-Weinbergin tasapainossa luonnonvalintaa lukuunottamatta, saadaan alleeleista muodostuvien eri tsygoottien genotyyppifrekvenssit
Metsägenetiikan sovellukset: Metsägenetiikan haasteet: geenit, geenivarat ja metsänjalostus
 Katri Kärkkäinen Matti Haapanen Metsägenetiikan sovellukset: Metsägenetiikan haasteet: geenit, geenivarat ja metsänjalostus Katri Kärkkäinen ja Matti Haapanen Metsäntutkimuslaitos Vantaan tutkimuskeskus
Katri Kärkkäinen Matti Haapanen Metsägenetiikan sovellukset: Metsägenetiikan haasteet: geenit, geenivarat ja metsänjalostus Katri Kärkkäinen ja Matti Haapanen Metsäntutkimuslaitos Vantaan tutkimuskeskus
Genomin evoluutio. Miten genomin koko ja rakenne muuttuvat ja miten sitä tutkitaan?
 Genomin evoluutio Miten genomin koko ja rakenne muuttuvat ja miten sitä tutkitaan? -duplikaatiot: geenien, geenin osien duplikaatiot, segmentaaliset, koko genomin duplikaatiot -duplikoituneiden geenien
Genomin evoluutio Miten genomin koko ja rakenne muuttuvat ja miten sitä tutkitaan? -duplikaatiot: geenien, geenin osien duplikaatiot, segmentaaliset, koko genomin duplikaatiot -duplikoituneiden geenien
OULUN YLIOPISTO, BIOLOGIAN LAITOS Puututkimus
 OULUN YLIOPISTO, BIOLOGIAN LAITOS Puututkimus Puu on yksilö, lajinsa edustaja, eliöyhteisönsä jäsen, esteettinen näky ja paljon muuta. Tässä harjoituksessa lähestytään puuta monipuolisesti ja harjoitellaan
OULUN YLIOPISTO, BIOLOGIAN LAITOS Puututkimus Puu on yksilö, lajinsa edustaja, eliöyhteisönsä jäsen, esteettinen näky ja paljon muuta. Tässä harjoituksessa lähestytään puuta monipuolisesti ja harjoitellaan
58131 Tietorakenteet ja algoritmit (syksy 2015) Toinen välikoe, malliratkaisut
 Toinen välikoe, malliratkaisut") Tietorakenteet ja algoritmit (syksy 0) Toinen välikoe, malliratkaisut. (a) Alussa puu näyttää tältä: Lisätään 4: 4 Tasapaino rikkoutuu solmussa. Tehdään kaksoiskierto ensin oikealle solmusta ja sitten
Tietorakenteet ja algoritmit (syksy 0) Toinen välikoe, malliratkaisut. (a) Alussa puu näyttää tältä: Lisätään 4: 4 Tasapaino rikkoutuu solmussa. Tehdään kaksoiskierto ensin oikealle solmusta ja sitten
DNA sukututkimuksen tukena
 Järvenpää 12,2,2019 Teuvo Ikonen teuvo.ikonen@welho.com DNA sukututkimuksen tukena DNA sukututkimuksessa (Peter Sjölund: Släktforska med DNA) tiesitkö, että olet kävelevä sukukirja? on kuin lukisit kirjaa
Järvenpää 12,2,2019 Teuvo Ikonen teuvo.ikonen@welho.com DNA sukututkimuksen tukena DNA sukututkimuksessa (Peter Sjölund: Släktforska med DNA) tiesitkö, että olet kävelevä sukukirja? on kuin lukisit kirjaa
Metsäpatologian laboratorio tuhotutkimuksen apuna. Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari
 Metsäpatologian laboratorio tuhotutkimuksen apuna Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari Metsäpuiden vaivat Metsäpuiden eloa ja terveyttä uhkaavat monet taudinaiheuttajat: Bioottiset taudinaiheuttajat
Metsäpatologian laboratorio tuhotutkimuksen apuna Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari Metsäpuiden vaivat Metsäpuiden eloa ja terveyttä uhkaavat monet taudinaiheuttajat: Bioottiset taudinaiheuttajat
Johdatus verkkoteoriaan 4. luento
 Johdatus verkkoteoriaan 4. luento 28.11.17 Viikolla 46 läpikäydyt käsitteet Viikolla 47 läpikäydyt käsitteet Verkko eli graafi, tasoverkko, solmut, välit, alueet, suunnatut verkot, isomorfiset verkot,
Johdatus verkkoteoriaan 4. luento 28.11.17 Viikolla 46 läpikäydyt käsitteet Viikolla 47 läpikäydyt käsitteet Verkko eli graafi, tasoverkko, solmut, välit, alueet, suunnatut verkot, isomorfiset verkot,
Graafit ja verkot. Joukko solmuja ja joukko järjestämättömiä solmupareja. eli haaroja. Joukko solmuja ja joukko järjestettyjä solmupareja eli kaaria
 Graafit ja verkot Suuntamaton graafi: eli haaroja Joukko solmuja ja joukko järjestämättömiä solmupareja Suunnattu graafi: Joukko solmuja ja joukko järjestettyjä solmupareja eli kaaria Haaran päätesolmut:
Graafit ja verkot Suuntamaton graafi: eli haaroja Joukko solmuja ja joukko järjestämättömiä solmupareja Suunnattu graafi: Joukko solmuja ja joukko järjestettyjä solmupareja eli kaaria Haaran päätesolmut:
DNA-testit. sukututkimuksessa Keravan kirjasto Paula Päivinen
 DNA-testit sukututkimuksessa 28.11.2017 Keravan kirjasto Paula Päivinen Solu tuma kromosomit 23 paria DNA Tumassa olevat kromosomit periytyvät jälkeläisille puoliksi isältä ja äidiltä Y-kromosomi periytyy
DNA-testit sukututkimuksessa 28.11.2017 Keravan kirjasto Paula Päivinen Solu tuma kromosomit 23 paria DNA Tumassa olevat kromosomit periytyvät jälkeläisille puoliksi isältä ja äidiltä Y-kromosomi periytyy
Perinnöllisyyden perusteita
 Perinnöllisyyden perusteita Eero Lukkari Tämä artikkeli kertoo perinnöllisyyden perusmekanismeista johdantona muille jalostus- ja terveysaiheisille artikkeleille. Koirien, kuten muidenkin eliöiden, perimä
Perinnöllisyyden perusteita Eero Lukkari Tämä artikkeli kertoo perinnöllisyyden perusmekanismeista johdantona muille jalostus- ja terveysaiheisille artikkeleille. Koirien, kuten muidenkin eliöiden, perimä
1. OHJAAMATON OPPIMINEN JA KLUSTEROINTI
 1. OHJAAMATON OPPIMINEN JA KLUSTEROINTI 1 1.1 Funktion optimointiin perustuvat klusterointialgoritmit Klusteroinnin onnistumista mittaavan funktion J optimointiin perustuvissa klusterointialgoritmeissä
1. OHJAAMATON OPPIMINEN JA KLUSTEROINTI 1 1.1 Funktion optimointiin perustuvat klusterointialgoritmit Klusteroinnin onnistumista mittaavan funktion J optimointiin perustuvissa klusterointialgoritmeissä
Ryhmät & uudet mahdollisuudet
 www.terrasolid.com Ryhmät & uudet mahdollisuudet Arttu Soininen 22.08.2017 Uudet mahdollisuudet ryhmien avulla Parempi maanpinnan yläpuolisten kohteiden luokittelu Maanpäällisten kohteiden luokittelu toimii
www.terrasolid.com Ryhmät & uudet mahdollisuudet Arttu Soininen 22.08.2017 Uudet mahdollisuudet ryhmien avulla Parempi maanpinnan yläpuolisten kohteiden luokittelu Maanpäällisten kohteiden luokittelu toimii
Algoritmit 2. Luento 12 Ke Timo Männikkö
 Algoritmit 2 Luento 12 Ke 26.4.2017 Timo Männikkö Luento 12 Rajoitehaku Kauppamatkustajan ongelma Lyhin virittävä puu Paikallinen etsintä Vaihtoalgoritmit Geneettiset algoritmit Simuloitu jäähdytys Algoritmit
Algoritmit 2 Luento 12 Ke 26.4.2017 Timo Männikkö Luento 12 Rajoitehaku Kauppamatkustajan ongelma Lyhin virittävä puu Paikallinen etsintä Vaihtoalgoritmit Geneettiset algoritmit Simuloitu jäähdytys Algoritmit
Algoritmit 2. Luento 6 Ke Timo Männikkö
 Algoritmit 2 Luento 6 Ke 29.3.2017 Timo Männikkö Luento 6 B-puun operaatiot B-puun muunnelmia Nelipuu Trie-rakenteet Standarditrie Pakattu trie Algoritmit 2 Kevät 2017 Luento 6 Ke 29.3.2017 2/31 B-puu
Algoritmit 2 Luento 6 Ke 29.3.2017 Timo Männikkö Luento 6 B-puun operaatiot B-puun muunnelmia Nelipuu Trie-rakenteet Standarditrie Pakattu trie Algoritmit 2 Kevät 2017 Luento 6 Ke 29.3.2017 2/31 B-puu
Genomin ilmentyminen Liisa Kauppi, Genomibiologian tutkimusohjelma
 Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Kombinatorinen optimointi
 Kombinatorinen optimointi Sallittujen pisteiden lukumäärä on äärellinen Periaatteessa ratkaisu löydetään käymällä läpi kaikki pisteet Käytännössä lukumäärä on niin suuri, että tämä on mahdotonta Usein
Kombinatorinen optimointi Sallittujen pisteiden lukumäärä on äärellinen Periaatteessa ratkaisu löydetään käymällä läpi kaikki pisteet Käytännössä lukumäärä on niin suuri, että tämä on mahdotonta Usein
Ratkaisu. Tulkitaan de Bruijnin jonon etsimiseksi aakkostossa S := {0, 1} sanapituudelle n = 4. Neljän pituisia sanoja on N = 2 n = 16 kpl.
 iskreetti matematiikka, syksy 00 arjoitus, ratkaisuista. seta 8 nollaa ja 8 ykköstä renkaaksi niin, että jokainen yhdistelmä 0000, 000,..., esiintyy täsmälleen kerran. Vihje: Tulkitse de ruijnin jonon
iskreetti matematiikka, syksy 00 arjoitus, ratkaisuista. seta 8 nollaa ja 8 ykköstä renkaaksi niin, että jokainen yhdistelmä 0000, 000,..., esiintyy täsmälleen kerran. Vihje: Tulkitse de ruijnin jonon
jens 1 matti Etäisyydet 1: 1.1 2: 1.4 3: 1.8 4: 2.0 5: 3.0 6: 3.6 7: 4.0 zetor
 T-1.81 Luonnollisten kielten tilastollinen käsittely Vastaukset 11, ti 8.4., 1:1-18: Klusterointi, Konekääntäminen. Versio 1. 1. Kuvaan 1 on piirretty klusteroinnit käyttäen annettuja algoritmeja. Sanojen
T-1.81 Luonnollisten kielten tilastollinen käsittely Vastaukset 11, ti 8.4., 1:1-18: Klusterointi, Konekääntäminen. Versio 1. 1. Kuvaan 1 on piirretty klusteroinnit käyttäen annettuja algoritmeja. Sanojen
805306A Johdatus monimuuttujamenetelmiin, 5 op
 monimuuttujamenetelmiin, 5 op syksy 2018 Matemaattisten tieteiden laitos K:n lähimmän naapurin menetelmä (K-Nearest neighbours) Tarkastellaan aluksi pientä (n = 9) kurjenmiekka-aineistoa, joka on seuraava:
monimuuttujamenetelmiin, 5 op syksy 2018 Matemaattisten tieteiden laitos K:n lähimmän naapurin menetelmä (K-Nearest neighbours) Tarkastellaan aluksi pientä (n = 9) kurjenmiekka-aineistoa, joka on seuraava:
j n j a b a c a d b c c d m j b a c a d a c b d c c j
 TEKNILLINEN KORKEAKOULU Tietoliikenne- ja tietoverkkotekniikan laitos S-38.115 Liikenneteorian perusteet, Kevät 2008 Demonstraatiot Luento 12 29.2.2008 D12/1 Tarkastellaan verkkoa, jossa on solmua ja linkkiä.
TEKNILLINEN KORKEAKOULU Tietoliikenne- ja tietoverkkotekniikan laitos S-38.115 Liikenneteorian perusteet, Kevät 2008 Demonstraatiot Luento 12 29.2.2008 D12/1 Tarkastellaan verkkoa, jossa on solmua ja linkkiä.
Algoritmi I kuvioiden ja niille johtavien ajourien erottelu. Metsätehon tuloskalvosarja 7a/2018 LIITE 1 Timo Melkas Kirsi Riekki Metsäteho Oy
 Algoritmi I kuvioiden ja niille johtavien ajourien erottelu Metsätehon tuloskalvosarja 7a/2018 LIITE 1 Timo Melkas Kirsi Riekki Metsäteho Oy Algoritmi I kuvioiden ja niille johtavien ajourien erottelu
Algoritmi I kuvioiden ja niille johtavien ajourien erottelu Metsätehon tuloskalvosarja 7a/2018 LIITE 1 Timo Melkas Kirsi Riekki Metsäteho Oy Algoritmi I kuvioiden ja niille johtavien ajourien erottelu
Luku 7. Verkkoalgoritmit. 7.1 Määritelmiä
 Luku 7 Verkkoalgoritmit Verkot soveltuvat monenlaisten ohjelmointiongelmien mallintamiseen. Tyypillinen esimerkki verkosta on tieverkosto, jonka rakenne muistuttaa luonnostaan verkkoa. Joskus taas verkko
Luku 7 Verkkoalgoritmit Verkot soveltuvat monenlaisten ohjelmointiongelmien mallintamiseen. Tyypillinen esimerkki verkosta on tieverkosto, jonka rakenne muistuttaa luonnostaan verkkoa. Joskus taas verkko
Sekvenssien rinnastus. Rinnastus: helppoa tai vaikeaa
 Sekvenssien rinnastus Rinnastus: helppoa tai vaikeaa Kaksi tai useampia (DNA tai proteiini) sekvenssejä: miten samankaltaisia sekvenssit ovat missä sekvenssikohdissa samankaltaisuutta esiintyy Kattava
Sekvenssien rinnastus Rinnastus: helppoa tai vaikeaa Kaksi tai useampia (DNA tai proteiini) sekvenssejä: miten samankaltaisia sekvenssit ovat missä sekvenssikohdissa samankaltaisuutta esiintyy Kattava
Uhanalaisuusluokat. Lajien uhanalaisuusarviointi Ulla-Maija Liukko, Arviointikoulutus lajien uhanalaisuuden arvioijille, 2.2.
 Uhanalaisuusluokat Lajien uhanalaisuusarviointi 2019 Ulla-Maija Liukko, Arviointikoulutus lajien uhanalaisuuden arvioijille, 2.2.2017 IUCN:n uhanalaisuusluokitus Uhanalaisuusarvioinnissa ja luokittelussa
Uhanalaisuusluokat Lajien uhanalaisuusarviointi 2019 Ulla-Maija Liukko, Arviointikoulutus lajien uhanalaisuuden arvioijille, 2.2.2017 IUCN:n uhanalaisuusluokitus Uhanalaisuusarvioinnissa ja luokittelussa
Luennon 3 oppimistavoitteet. Solulajit PUUSOLUT. Luennon 3 oppimistavoitteet. Puu-19.210 Puun rakenne ja kemia
 Solulajit Puu-19.210 Puun rakenne ja kemia Luennon 3 oppimistavoitteet Osaat luokitella puusolut ja ymmärrät niiden tehtävät ja sijainnin puusolukossa. Tunnistat havupuiden ja lehtipuiden solukot mikroskooppikuvista.
Solulajit Puu-19.210 Puun rakenne ja kemia Luennon 3 oppimistavoitteet Osaat luokitella puusolut ja ymmärrät niiden tehtävät ja sijainnin puusolukossa. Tunnistat havupuiden ja lehtipuiden solukot mikroskooppikuvista.
SSL syysseminaari 29.10.2013 Juha Hyssälä
 SSL syysseminaari 29.10.2013 Juha Hyssälä Lääketieteellisessä tutkimuksessa on perinteisesti käytetty elinaika-analyysissä Coxin suhteellisen vaaran mallia ja/tai tämän johdannaisia. Kyseinen malli kuitenkin
SSL syysseminaari 29.10.2013 Juha Hyssälä Lääketieteellisessä tutkimuksessa on perinteisesti käytetty elinaika-analyysissä Coxin suhteellisen vaaran mallia ja/tai tämän johdannaisia. Kyseinen malli kuitenkin
Avainsanojen poimiminen Eeva Ahonen
 Avainsanojen poimiminen 5.10.2004 Eeva Ahonen Sisältö Avainsanat Menetelmät C4.5 päätöspuut GenEx algoritmi Bayes malli Testit Tulokset Avainsanat Tiivistä tietoa dokumentin sisällöstä ihmislukijalle hakukoneelle
Avainsanojen poimiminen 5.10.2004 Eeva Ahonen Sisältö Avainsanat Menetelmät C4.5 päätöspuut GenEx algoritmi Bayes malli Testit Tulokset Avainsanat Tiivistä tietoa dokumentin sisällöstä ihmislukijalle hakukoneelle
Etsintä verkosta (Searching from the Web) T Datasta tietoon Heikki Mannila, Jouni Seppänen
 T Datasta tietoon Heikki Mannila, Jouni Seppänen") Etsintä verkosta (Searching from the Web) T-61.2010 Datasta tietoon Heikki Mannila, Jouni Seppänen 12.12.2007 Webin lyhyt historia http://info.cern.ch/proposal.html http://browser.arachne.cz/screen/
Etsintä verkosta (Searching from the Web) T-61.2010 Datasta tietoon Heikki Mannila, Jouni Seppänen 12.12.2007 Webin lyhyt historia http://info.cern.ch/proposal.html http://browser.arachne.cz/screen/
Laatua kuvaavien kasvumallien kehittäminen. Annikki Mäkelä, Anu Kantola, Harri Mäkinen HY Metsäekologian laitos, Metla
 Laatua kuvaavien kasvumallien kehittäminen Annikki Mäkelä, Anu Kantola, Harri Mäkinen HY Metsäekologian laitos, Metla Taustaa» Kasvumallit antavat puustoennusteen kiertoaikana, kun tunnetaan» kasvupaikkatiedot»
Laatua kuvaavien kasvumallien kehittäminen Annikki Mäkelä, Anu Kantola, Harri Mäkinen HY Metsäekologian laitos, Metla Taustaa» Kasvumallit antavat puustoennusteen kiertoaikana, kun tunnetaan» kasvupaikkatiedot»
DBN Mitä sillä tekee? Dynaamisten Bayes-verkkojen määrittely aikasarja-analyysissä Janne Toivola jtoivola@iki.fi
 DBN Mitä sillä tekee? Dynaamisten Bayes-verkkojen määrittely aikasarja-analyysissä Janne Toivola jtoivola@iki.fi Historiaa Bayesin kaavan hyödyntäminen BN-ohjelmistoja ollut ennenkin Tanskalaisten Hugin
DBN Mitä sillä tekee? Dynaamisten Bayes-verkkojen määrittely aikasarja-analyysissä Janne Toivola jtoivola@iki.fi Historiaa Bayesin kaavan hyödyntäminen BN-ohjelmistoja ollut ennenkin Tanskalaisten Hugin
Männyn laaturajojen integrointi runkokäyrän ennustamisessa. Laura Koskela Tampereen yliopisto 9.6.2003
 Männyn laaturajojen integrointi runkokäyrän ennustamisessa Laura Koskela Tampereen yliopisto 9.6.2003 Johdantoa Pohjoismaisen käytännön mukaan rungot katkaistaan tukeiksi jo metsässä. Katkonnan ohjauksessa
Männyn laaturajojen integrointi runkokäyrän ennustamisessa Laura Koskela Tampereen yliopisto 9.6.2003 Johdantoa Pohjoismaisen käytännön mukaan rungot katkaistaan tukeiksi jo metsässä. Katkonnan ohjauksessa
Molekyylipopulaatiogenetiikka
 Molekyylipopulaatiogenetiikka Hedrick 2005, kappale 8, pp. 452-462, 428-449 Demografian ja valinnan vaikutukset koalesenssipuihin Valinnan havaitseminen TajimanD Divergenssin ja polymorfismin vertailu
Molekyylipopulaatiogenetiikka Hedrick 2005, kappale 8, pp. 452-462, 428-449 Demografian ja valinnan vaikutukset koalesenssipuihin Valinnan havaitseminen TajimanD Divergenssin ja polymorfismin vertailu
Laskennallinen menetelmä puun biomassan ja oksien kokojakauman määrittämiseen laserkeilausdatasta
 Laskennallinen menetelmä puun biomassan ja oksien kokojakauman määrittämiseen laserkeilausdatasta Pasi Raumonen, Mikko Kaasalainen ja Markku Åkerblom Tampereen teknillinen ylipisto, Matematiikan laitos
Laskennallinen menetelmä puun biomassan ja oksien kokojakauman määrittämiseen laserkeilausdatasta Pasi Raumonen, Mikko Kaasalainen ja Markku Åkerblom Tampereen teknillinen ylipisto, Matematiikan laitos
Lisääntyminen. BI1 Elämä ja evoluutio Leena kangas-järviluoma
 Lisääntyminen BI1 Elämä ja evoluutio Leena kangas-järviluoma säilyä hengissä ja lisääntyä kaksi tapaa lisääntyä suvuton suvullinen suvuttomassa lisääntymisessä uusi yksilö syntyy ilman sukusoluja suvullisessa
Lisääntyminen BI1 Elämä ja evoluutio Leena kangas-järviluoma säilyä hengissä ja lisääntyä kaksi tapaa lisääntyä suvuton suvullinen suvuttomassa lisääntymisessä uusi yksilö syntyy ilman sukusoluja suvullisessa
Datatähti 2019 loppu
 Datatähti 2019 loppu task type time limit memory limit A Summa standard 1.00 s 512 MB B Bittijono standard 1.00 s 512 MB C Auringonlasku standard 1.00 s 512 MB D Binääripuu standard 1.00 s 512 MB E Funktio
Datatähti 2019 loppu task type time limit memory limit A Summa standard 1.00 s 512 MB B Bittijono standard 1.00 s 512 MB C Auringonlasku standard 1.00 s 512 MB D Binääripuu standard 1.00 s 512 MB E Funktio
58131 Tietorakenteet (kevät 2009) Harjoitus 6, ratkaisuja (Antti Laaksonen)
 Harjoitus 6, ratkaisuja (Antti Laaksonen)") 58131 Tietorakenteet (kevät 2009) Harjoitus 6, ratkaisuja (Antti Laaksonen) 1. Avaimet 1, 2, 3 ja 4 mahtuvat samaan lehtisolmuun. Tässä tapauksessa puussa on vain yksi solmu, joka on samaan aikaan juurisolmu
58131 Tietorakenteet (kevät 2009) Harjoitus 6, ratkaisuja (Antti Laaksonen) 1. Avaimet 1, 2, 3 ja 4 mahtuvat samaan lehtisolmuun. Tässä tapauksessa puussa on vain yksi solmu, joka on samaan aikaan juurisolmu
811312A Tietorakenteet ja algoritmit 2015-2016. V Verkkojen algoritmeja Osa 2 : Kruskalin ja Dijkstran algoritmit
 811312A Tietorakenteet ja algoritmit 2015-2016 V Verkkojen algoritmeja Osa 2 : Kruskalin ja Dijkstran algoritmit Sisältö 1. Johdanto 2. Leveyshaku 3. Syvyyshaku 4. Kruskalin algoritmi 5. Dijkstran algoritmi
811312A Tietorakenteet ja algoritmit 2015-2016 V Verkkojen algoritmeja Osa 2 : Kruskalin ja Dijkstran algoritmit Sisältö 1. Johdanto 2. Leveyshaku 3. Syvyyshaku 4. Kruskalin algoritmi 5. Dijkstran algoritmi
Sukukokous TERVETULOA! Järvisydän, Rantasalmi
 Sukukokous 1.7.2017 TERVETULOA! Järvisydän, Rantasalmi Kansasten DNA-sukututkimus TAUSTAA Mitä on DNA-sukututkimus? Vahvistaa perinteisin sukututkimusmenetelmin saatuja tuloksia. Tutkitaan henkilön DNA-näytettä.
Sukukokous 1.7.2017 TERVETULOA! Järvisydän, Rantasalmi Kansasten DNA-sukututkimus TAUSTAA Mitä on DNA-sukututkimus? Vahvistaa perinteisin sukututkimusmenetelmin saatuja tuloksia. Tutkitaan henkilön DNA-näytettä.
Algoritmit 2. Luento 11 Ti Timo Männikkö
 Algoritmit 2 Luento 11 Ti 24.4.2018 Timo Männikkö Luento 11 Rajoitehaku Kapsäkkiongelma Kauppamatkustajan ongelma Paikallinen etsintä Lyhin virittävä puu Vaihtoalgoritmit Algoritmit 2 Kevät 2018 Luento
Algoritmit 2 Luento 11 Ti 24.4.2018 Timo Männikkö Luento 11 Rajoitehaku Kapsäkkiongelma Kauppamatkustajan ongelma Paikallinen etsintä Lyhin virittävä puu Vaihtoalgoritmit Algoritmit 2 Kevät 2018 Luento
DENDROKRONOLOGIAN LABORATORIO METSÄTIETEIDEN OSASTO LUONNONTIETEIDEN JA METSÄ TIETEIDEN TIEDEKUNTA ITÄ-SUOMEN YLIOPISTO, JOENSUU
 DENDROKRONOLOGIAN LABORATORIO METSÄTIETEIDEN OSASTO LUONNONTIETEIDEN JA METSÄ TIETEIDEN TIEDEKUNTA ITÄ-SUOMEN YLIOPISTO, JOENSUU Kallonlahden hylky ja siitä tutkitut kuusi dendrokronologista ajoitusnäytettä.
DENDROKRONOLOGIAN LABORATORIO METSÄTIETEIDEN OSASTO LUONNONTIETEIDEN JA METSÄ TIETEIDEN TIEDEKUNTA ITÄ-SUOMEN YLIOPISTO, JOENSUU Kallonlahden hylky ja siitä tutkitut kuusi dendrokronologista ajoitusnäytettä.
Geenikartoitusmenetelmät. Kytkentäanalyysin teoriaa. Suurimman uskottavuuden menetelmä ML (maximum likelihood) Uskottavuusfunktio: koko aineisto
 Uskottavuusfunktio: koko aineisto") Kytkentäanalyysin teoriaa Pyritään selvittämään tiettyyn ominaisuuteen vaikuttavien eenien paikka enomissa Perustavoite: löytää markkerilokus jonka alleelit ja tutkittava ominaisuus (esim. sairaus) periytyvät
Kytkentäanalyysin teoriaa Pyritään selvittämään tiettyyn ominaisuuteen vaikuttavien eenien paikka enomissa Perustavoite: löytää markkerilokus jonka alleelit ja tutkittava ominaisuus (esim. sairaus) periytyvät
Steven Kelly & Mia+Janne
 Luomisoppi evoluutio Steven Kelly & Mia+Janne Tämä ei ole väittely! Pidetään kiinni yhteisestä uskosta: Alussa Jumala loi Se, että on Luoja, ratkaisee paljon: käytetään sitä rohkeasti apologiassa Eri mielipiteitä
Luomisoppi evoluutio Steven Kelly & Mia+Janne Tämä ei ole väittely! Pidetään kiinni yhteisestä uskosta: Alussa Jumala loi Se, että on Luoja, ratkaisee paljon: käytetään sitä rohkeasti apologiassa Eri mielipiteitä
GA & robot path planning. Janne Haapsaari AUTO Geneettiset algoritmit
 GA & robot path planning Janne Haapsaari AUTO3070 - Geneettiset algoritmit GA robotiikassa Sovelluksia liikkeen optimoinnissa: * eri vapausasteisten robottien liikeratojen optimointi * autonomisten robottien
GA & robot path planning Janne Haapsaari AUTO3070 - Geneettiset algoritmit GA robotiikassa Sovelluksia liikkeen optimoinnissa: * eri vapausasteisten robottien liikeratojen optimointi * autonomisten robottien
Bayesin pelit. Kalle Siukola. MS-E2142 Optimointiopin seminaari: Peliteoria ja tekoäly
 Bayesin pelit Kalle Siukola MS-E2142 Optimointiopin seminaari: Peliteoria ja tekoäly 12.10.2016 Toistetun pelin esittäminen automaatin avulla Ekstensiivisen muodon puu on tehoton esitystapa, jos peliä
Bayesin pelit Kalle Siukola MS-E2142 Optimointiopin seminaari: Peliteoria ja tekoäly 12.10.2016 Toistetun pelin esittäminen automaatin avulla Ekstensiivisen muodon puu on tehoton esitystapa, jos peliä
ORMS2020 Päätöksenteko epävarmuuden vallitessa Syksy 2008. päätöspuiden avulla tarkastellaan vasta seuraavissa harjoituksissa.
 ORMS00 Päätöksenteko epävarmuuden vallitessa Syksy 008 Harjoitus Ratkaisuehdotuksia Nämä harjoitukset liittyvät päätöspuiden rakentamiseen: varsinaista päätöksentekoa päätöspuiden avulla tarkastellaan
ORMS00 Päätöksenteko epävarmuuden vallitessa Syksy 008 Harjoitus Ratkaisuehdotuksia Nämä harjoitukset liittyvät päätöspuiden rakentamiseen: varsinaista päätöksentekoa päätöspuiden avulla tarkastellaan
KASVIKUNTA kl 2016
 526128 KASVIKUNTA kl 2016 Luennot klo 10-12 salissa 6602 11.4. Ma johdanto kasvikuntaan 12.4. Ti Chlorophyta 14.4. To Streptophyta 15.4. Pe vanhimmat alkiolliset kasvit 18.4. Ma maksasammalet 19.4. Ti
526128 KASVIKUNTA kl 2016 Luennot klo 10-12 salissa 6602 11.4. Ma johdanto kasvikuntaan 12.4. Ti Chlorophyta 14.4. To Streptophyta 15.4. Pe vanhimmat alkiolliset kasvit 18.4. Ma maksasammalet 19.4. Ti
Johdatus graafiteoriaan
 Johdatus graafiteoriaan Syksy 2017 Lauri Hella Tampereen yliopisto Luonnontieteiden tiedekunta 126 Luku 3 Puut 3.1 Puu 3.2 Virittävä puu 3.3 Virittävän puun konstruointi 3.4 Minimaalinen virittävä puu
Johdatus graafiteoriaan Syksy 2017 Lauri Hella Tampereen yliopisto Luonnontieteiden tiedekunta 126 Luku 3 Puut 3.1 Puu 3.2 Virittävä puu 3.3 Virittävän puun konstruointi 3.4 Minimaalinen virittävä puu
T Luonnollisen kielen tilastollinen käsittely Vastaukset 3, ti , 8:30-10:00 Kollokaatiot, Versio 1.1
 T-61.281 Luonnollisen kielen tilastollinen käsittely Vastaukset 3, ti 10.2.2004, 8:30-10:00 Kollokaatiot, Versio 1.1 1. Lasketaan ensin tulokset sanaparille valkoinen, talo käsin: Frekvenssimenetelmä:
T-61.281 Luonnollisen kielen tilastollinen käsittely Vastaukset 3, ti 10.2.2004, 8:30-10:00 Kollokaatiot, Versio 1.1 1. Lasketaan ensin tulokset sanaparille valkoinen, talo käsin: Frekvenssimenetelmä:
Arkkitehtuurien tutkimus Outi Räihä. OHJ-3200 Ohjelmistoarkkitehtuurit. Darwin-projekti. Johdanto
 OHJ-3200 Ohjelmistoarkkitehtuurit 1 Arkkitehtuurien tutkimus Outi Räihä 2 Darwin-projekti Darwin-projekti: Akatemian rahoitus 2009-2011 Arkkitehtuurisuunnittelu etsintäongelmana Geneettiset algoritmit
OHJ-3200 Ohjelmistoarkkitehtuurit 1 Arkkitehtuurien tutkimus Outi Räihä 2 Darwin-projekti Darwin-projekti: Akatemian rahoitus 2009-2011 Arkkitehtuurisuunnittelu etsintäongelmana Geneettiset algoritmit
13 Lyhimmät painotetut polut
 TIE-20100 Tietorakenteet ja algoritmit 297 13 Lyhimmät painotetut polut BFS löytää lyhimmän polun lähtösolmusta graafin saavutettaviin solmuihin. Se ei kuitenkaan enää suoriudu tehtävästä, jos kaarien
TIE-20100 Tietorakenteet ja algoritmit 297 13 Lyhimmät painotetut polut BFS löytää lyhimmän polun lähtösolmusta graafin saavutettaviin solmuihin. Se ei kuitenkaan enää suoriudu tehtävästä, jos kaarien
3. Hakupuut. B-puu on hakupuun laji, joka sopii mm. tietokantasovelluksiin, joissa rakenne on talletettu kiintolevylle eikä keskusmuistiin.
 3. Hakupuut Hakupuu on listaa tehokkaampi dynaamisen joukon toteutus. Erityisesti suurilla tietomäärillä hakupuu kannattaa tasapainottaa, jolloin päivitysoperaatioista tulee hankalampia toteuttaa mutta
3. Hakupuut Hakupuu on listaa tehokkaampi dynaamisen joukon toteutus. Erityisesti suurilla tietomäärillä hakupuu kannattaa tasapainottaa, jolloin päivitysoperaatioista tulee hankalampia toteuttaa mutta
Algoritmit 2. Luento 13 Ti Timo Männikkö
 Algoritmit 2 Luento 13 Ti 30.4.2019 Timo Männikkö Luento 13 Simuloitu jäähdytys Merkkijonon sovitus Horspoolin algoritmi Ositus ja rekursio Rekursion toteutus Algoritmit 2 Kevät 2019 Luento 13 Ti 30.4.2019
Algoritmit 2 Luento 13 Ti 30.4.2019 Timo Männikkö Luento 13 Simuloitu jäähdytys Merkkijonon sovitus Horspoolin algoritmi Ositus ja rekursio Rekursion toteutus Algoritmit 2 Kevät 2019 Luento 13 Ti 30.4.2019
Bayesilainen päätöksenteko / Bayesian decision theory
 Bayesilainen päätöksenteko / Bayesian decision theory Todennäköisyysteoria voidaan perustella ilman päätösteoriaa, mutta vasta päätösteorian avulla siitä on oikeasti hyötyä Todennäköisyyteoriassa tavoitteena
Bayesilainen päätöksenteko / Bayesian decision theory Todennäköisyysteoria voidaan perustella ilman päätösteoriaa, mutta vasta päätösteorian avulla siitä on oikeasti hyötyä Todennäköisyyteoriassa tavoitteena
Tekijä Pitkä matematiikka On osoitettava, että jana DE sivun AB kanssa yhdensuuntainen ja sen pituus on 4 5
 Tekijä Pitkä matematiikka 6..06 8 On osoitettava, että jana DE sivun AB kanssa yhdensuuntainen ja sen pituus on 5 sivun AB pituudesta. Pitää siis osoittaa, että DE = AB. 5 Muodostetaan vektori DE. DE =
Tekijä Pitkä matematiikka 6..06 8 On osoitettava, että jana DE sivun AB kanssa yhdensuuntainen ja sen pituus on 5 sivun AB pituudesta. Pitää siis osoittaa, että DE = AB. 5 Muodostetaan vektori DE. DE =
Pinot, jonot, yleisemmin sekvenssit: kokoelma peräkkäisiä alkioita (lineaarinen järjestys) Yleisempi tilanne: alkioiden hierarkia
 Yleisempi tilanne: alkioiden hierarkia") Pinot, jonot, yleisemmin sekvenssit: kokoelma peräkkäisiä alkioita (lineaarinen järjestys) Yleisempi tilanne: alkioiden hierarkia Kukin alkio (viite) talletettuna solmuun (node) vastaa paikan käsitettä
Pinot, jonot, yleisemmin sekvenssit: kokoelma peräkkäisiä alkioita (lineaarinen järjestys) Yleisempi tilanne: alkioiden hierarkia Kukin alkio (viite) talletettuna solmuun (node) vastaa paikan käsitettä
Taistelu tyvitervastautia vastaan tutkimustieto laboratoriosta käytäntöön
 Taistelu tyvitervastautia vastaan tutkimustieto laboratoriosta käytäntöön Metsätieteen päivä 19.10.2016 Tuula Piri Luonnonvarakeskus 1 26.10.2016 Männyn tyvitervastauti Aiheuttaja männynjuurikääpä (Heterobasidion
Taistelu tyvitervastautia vastaan tutkimustieto laboratoriosta käytäntöön Metsätieteen päivä 19.10.2016 Tuula Piri Luonnonvarakeskus 1 26.10.2016 Männyn tyvitervastauti Aiheuttaja männynjuurikääpä (Heterobasidion
Biopankit miksi ja millä ehdoilla?
 Suomalaisen Tiedeakatemian 100 v-symposium, Helsinki 4.9.2008 Biopankit miksi ja millä ehdoilla? Juha Kere Karolinska Institutet, Stockholm, Sverige ja Helsingin yliopisto Tautien tutkimus Geeni/ valkuaisaine
Suomalaisen Tiedeakatemian 100 v-symposium, Helsinki 4.9.2008 Biopankit miksi ja millä ehdoilla? Juha Kere Karolinska Institutet, Stockholm, Sverige ja Helsingin yliopisto Tautien tutkimus Geeni/ valkuaisaine
Algoritmit 1. Luento 8 Ke Timo Männikkö
 Algoritmit 1 Luento 8 Ke 1.2.2017 Timo Männikkö Luento 8 Järjestetty binääripuu Solmujen läpikäynti Binääripuun korkeus Binääripuun tasapainottaminen Graafit ja verkot Verkon lyhimmät polut Fordin ja Fulkersonin
Algoritmit 1 Luento 8 Ke 1.2.2017 Timo Männikkö Luento 8 Järjestetty binääripuu Solmujen läpikäynti Binääripuun korkeus Binääripuun tasapainottaminen Graafit ja verkot Verkon lyhimmät polut Fordin ja Fulkersonin
Pikalajittelu: valitaan ns. pivot-alkio esim. pivot = oikeanpuoleisin
 Pikalajittelu: valitaan ns. pivot-alkio esim. pivot = oikeanpuoleisin jaetaan muut alkiot kahteen ryhmään: L: alkiot, jotka eivät suurempia kuin pivot G : alkiot, jotka suurempia kuin pivot 6 1 4 3 7 2
Pikalajittelu: valitaan ns. pivot-alkio esim. pivot = oikeanpuoleisin jaetaan muut alkiot kahteen ryhmään: L: alkiot, jotka eivät suurempia kuin pivot G : alkiot, jotka suurempia kuin pivot 6 1 4 3 7 2
Evoluutio ja luominen. Mian tekemä esitys Jannen esittämänä
 Evoluutio ja luominen Mian tekemä esitys Jannen esittämänä Väite: tiedemiehet ovat todistaneet evoluutioteorian todeksi Evoluutioteorialla tässä tarkoitan teoriaa, jonka mukaan kaikki elollinen on kehittynyt
Evoluutio ja luominen Mian tekemä esitys Jannen esittämänä Väite: tiedemiehet ovat todistaneet evoluutioteorian todeksi Evoluutioteorialla tässä tarkoitan teoriaa, jonka mukaan kaikki elollinen on kehittynyt
TIEA241 Automaatit ja kieliopit, syksy Antti-Juhani Kaijanaho. 30. marraskuuta 2015
 TIEA241 Automaatit ja kieliopit, syksy 2015 Antti-Juhani Kaijanaho TIETOTEKNIIKAN LAITOS 30. marraskuuta 2015 Sisällys t Väitöstilaisuus 4.12.2015 kello 12 vanhassa juhlasalissa S212 saa tulla 2 demoruksia
TIEA241 Automaatit ja kieliopit, syksy 2015 Antti-Juhani Kaijanaho TIETOTEKNIIKAN LAITOS 30. marraskuuta 2015 Sisällys t Väitöstilaisuus 4.12.2015 kello 12 vanhassa juhlasalissa S212 saa tulla 2 demoruksia
Peittyvä periytyminen. Potilasopas. Kuvat: Rebecca J Kent www.rebeccajkent.com rebecca@rebeccajkent.com
 12 Peittyvä periytyminen Muokattu allamainittujen instanssien julkaisemista vihkosista, heidän laatustandardiensa mukaan: Guy's and St Thomas' Hospital, London, United Kingdom; and the London IDEAS Genetic
12 Peittyvä periytyminen Muokattu allamainittujen instanssien julkaisemista vihkosista, heidän laatustandardiensa mukaan: Guy's and St Thomas' Hospital, London, United Kingdom; and the London IDEAS Genetic
Avainsanat: perimä dna rna 5`-ja 3`-päät replikaatio polymeraasientsyymi eksoni introni promoottori tehostajajakso silmukointi mutaatio
 Avainsanat: perimä dna rna 5`-ja 3`-päät replikaatio polymeraasientsyymi eksoni introni promoottori tehostajajakso silmukointi mutaatio Perinnöllinen informaatio sijaitsee dna:ssa eli deoksiribonukleiinihapossa
Avainsanat: perimä dna rna 5`-ja 3`-päät replikaatio polymeraasientsyymi eksoni introni promoottori tehostajajakso silmukointi mutaatio Perinnöllinen informaatio sijaitsee dna:ssa eli deoksiribonukleiinihapossa
Bioteknologian perustyökaluja
 Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
Harjoitus 6 ( )
") Harjoitus 6 (21.4.2015) Tehtävä 1 Määritelmän (ks. luentomoniste s. 109) mukaan yleisen, muotoa min f(x) s. t. g(x) 0 h(x) = 0 x X olevan optimointitehtävän Lagrangen duaali on missä max θ(u, v) s. t.
Harjoitus 6 (21.4.2015) Tehtävä 1 Määritelmän (ks. luentomoniste s. 109) mukaan yleisen, muotoa min f(x) s. t. g(x) 0 h(x) = 0 x X olevan optimointitehtävän Lagrangen duaali on missä max θ(u, v) s. t.
x 7 3 4x x 7 4x 3 ( 7 4)x 3 : ( 7 4), 7 4 1,35 < ln x + 1 = ln ln u 2 3u 4 = 0 (u 4)(u + 1) = 0 ei ratkaisua
x 3 : ( 7 4), 7 4 1,35 < ln x + 1 = ln ln u 2 3u 4 = 0 (u 4)(u + 1) = 0 ei ratkaisua") Mallivastaukset - Harjoituskoe E E a) x 7 3 4x x 7 4x 3 ( 7 4)x 3 : ( 7 4), 7 4,35 < 0 x 3 7 4 b) 0 / x + dx = 0 ln x + = ln + ln 0 + = ln 0 Vastaus: ln c) x 4 3x 4 = 0 Sijoitetaan x = u Tulon nollasääntö
Mallivastaukset - Harjoituskoe E E a) x 7 3 4x x 7 4x 3 ( 7 4)x 3 : ( 7 4), 7 4,35 < 0 x 3 7 4 b) 0 / x + dx = 0 ln x + = ln + ln 0 + = ln 0 Vastaus: ln c) x 4 3x 4 = 0 Sijoitetaan x = u Tulon nollasääntö
805306A Johdatus monimuuttujamenetelmiin, 5 op
 monimuuttujamenetelmiin, 5 op syksy 2018 Matemaattisten tieteiden laitos Lineaarinen erotteluanalyysi (LDA, Linear discriminant analysis) Erotteluanalyysin avulla pyritään muodostamaan selittävistä muuttujista
monimuuttujamenetelmiin, 5 op syksy 2018 Matemaattisten tieteiden laitos Lineaarinen erotteluanalyysi (LDA, Linear discriminant analysis) Erotteluanalyysin avulla pyritään muodostamaan selittävistä muuttujista
Nollasummapelit ja bayesilaiset pelit
 Nollasummapelit ja bayesilaiset pelit Kristian Ovaska HELSINGIN YLIOPISTO Tietojenkäsittelytieteen laitos Seminaari: Peliteoria Helsinki 18. syyskuuta 2006 Sisältö 1 Johdanto 1 2 Nollasummapelit 1 2.1
Nollasummapelit ja bayesilaiset pelit Kristian Ovaska HELSINGIN YLIOPISTO Tietojenkäsittelytieteen laitos Seminaari: Peliteoria Helsinki 18. syyskuuta 2006 Sisältö 1 Johdanto 1 2 Nollasummapelit 1 2.1
Virkaanastujaisesitelmä Anneli Hoikkala. Evoluutotutkimus: ekologiaa ja molekyyligenetiikkaa
 Virkaanastujaisesitelmä 26.10.2003 Anneli Hoikkala Evoluutotutkimus: ekologiaa ja molekyyligenetiikkaa Evoluutiotutkimus on ollut kauan hyvin kiistelty tieteen ala. Evoluutioteoria on tuonut uudenlaisen
Virkaanastujaisesitelmä 26.10.2003 Anneli Hoikkala Evoluutotutkimus: ekologiaa ja molekyyligenetiikkaa Evoluutiotutkimus on ollut kauan hyvin kiistelty tieteen ala. Evoluutioteoria on tuonut uudenlaisen
Mediaanisuodattimet. Tähän asti käsitellyt suodattimet ovat olleet lineaarisia. Niille on tyypillistä, että. niiden ominaisuudet tunnetaan hyvin
 Mediaanisuodattimet Tähän asti käsitellyt suodattimet ovat olleet lineaarisia. Niille on tyypillistä, että niiden ominaisuudet tunnetaan hyvin niiden analysointiin on olemassa vakiintuneita menetelmiä
Mediaanisuodattimet Tähän asti käsitellyt suodattimet ovat olleet lineaarisia. Niille on tyypillistä, että niiden ominaisuudet tunnetaan hyvin niiden analysointiin on olemassa vakiintuneita menetelmiä
Meripihka. Trilobiitti. 1. Fossiilit. Hominidin kallo. Kivettynyt metsä. Ramses Suuri. Jäätynyt mammutti. Jäämies
 Meripihka 1. Fossiilit Trilobiitti Hominidin kallo Ramses Suuri Kivettynyt metsä Jäämies Jäätynyt mammutti Fossiili = aiemmalta geologiselta kaudelta peräisin oleva eliön jäänne (sanakirjan mukaan myös
Meripihka 1. Fossiilit Trilobiitti Hominidin kallo Ramses Suuri Kivettynyt metsä Jäämies Jäätynyt mammutti Fossiili = aiemmalta geologiselta kaudelta peräisin oleva eliön jäänne (sanakirjan mukaan myös
Jakaumamallit MELA2009:ssä. MELA käyttäjäpäivä Kari Härkönen
 Jakaumamallit MELA29:ssä MELA käyttäjäpäivä 11.11.29 Kari Härkönen Metsäntutkimuslaitos Skogsforskningsinstitutet Finnish Forest Research Institute www.metla.fi Aineistonmuodostuksessa useita vaihtoehtoisia
Jakaumamallit MELA29:ssä MELA käyttäjäpäivä 11.11.29 Kari Härkönen Metsäntutkimuslaitos Skogsforskningsinstitutet Finnish Forest Research Institute www.metla.fi Aineistonmuodostuksessa useita vaihtoehtoisia
Proteiinien kontaktiresidyjen ennustaminen. Tuomo Hartonen Teoreettisen fysiikan syventävien opintojen seminaari
 Proteiinien kontaktiresidyjen ennustaminen Tuomo Hartonen Teoreettisen fysiikan syventävien opintojen seminaari 13.12.12 Terminologiaa Aminohappo = proteiinien rakennuspalikka, luonto käyttää 20 erilaista
Proteiinien kontaktiresidyjen ennustaminen Tuomo Hartonen Teoreettisen fysiikan syventävien opintojen seminaari 13.12.12 Terminologiaa Aminohappo = proteiinien rakennuspalikka, luonto käyttää 20 erilaista
Epigeneettinen säätely ja genomin leimautuminen. Tiina Immonen BLL Biokemia ja kehitysbiologia
 Epigeneettinen säätely ja genomin leimautuminen Tiina Immonen BLL Biokemia ja kehitysbiologia 21.1.2014 Epigeneettinen säätely Epigenetic: may be used for anything to do with development, but nowadays
Epigeneettinen säätely ja genomin leimautuminen Tiina Immonen BLL Biokemia ja kehitysbiologia 21.1.2014 Epigeneettinen säätely Epigenetic: may be used for anything to do with development, but nowadays
Evoluutio. BI Elämä ja evoluutio Leena Kangas-Järviluoma
 Evoluutio BI Elämä ja evoluutio Leena Kangas-Järviluoma 1 Evoluutio lajinkehitystä, jossa eliölajit muuttuvat ja niistä voi kehittyä uusia lajeja on jatkunut elämän synnystä saakka, sillä ei ole päämäärää
Evoluutio BI Elämä ja evoluutio Leena Kangas-Järviluoma 1 Evoluutio lajinkehitystä, jossa eliölajit muuttuvat ja niistä voi kehittyä uusia lajeja on jatkunut elämän synnystä saakka, sillä ei ole päämäärää
Relevanttien sivujen etsintä verkosta: satunnaiskulut verkossa Linkkikeskukset ja auktoriteetit (hubs and authorities) -algoritmi
 -algoritmi") Kurssin loppuosa Diskreettejä menetelmiä laajojen 0-1 datajoukkojen analyysiin Kattavat joukot ja niiden etsintä tasoittaisella algoritmilla Relevanttien sivujen etsintä verkosta: satunnaiskulut verkossa
Kurssin loppuosa Diskreettejä menetelmiä laajojen 0-1 datajoukkojen analyysiin Kattavat joukot ja niiden etsintä tasoittaisella algoritmilla Relevanttien sivujen etsintä verkosta: satunnaiskulut verkossa
HMM ja geenien etsintä
 Kuten makovin mallien yhteydessä, niin HMM halutulla topologialla voidaan opettaa tunnistamaan geenejä. Ohessa eäs geenitunnistukseen käytetty topologia, joka tunnistaa ihmisen geenit (5 -> 3 ). Edellä
Kuten makovin mallien yhteydessä, niin HMM halutulla topologialla voidaan opettaa tunnistamaan geenejä. Ohessa eäs geenitunnistukseen käytetty topologia, joka tunnistaa ihmisen geenit (5 -> 3 ). Edellä
Mikäli huomaat virheen tai on kysyttävää liittyen malleihin, lähetä viesti osoitteeseen
 Mikäli huomaat virheen tai on kysyttävää liittyen malleihin, lähetä viesti osoitteeseen anton.mallasto@aalto.fi. 1. 2. Muista. Ryhmän G aliryhmä H on normaali aliryhmä, jos ah = Ha kaikilla a G. Toisin
Mikäli huomaat virheen tai on kysyttävää liittyen malleihin, lähetä viesti osoitteeseen anton.mallasto@aalto.fi. 1. 2. Muista. Ryhmän G aliryhmä H on normaali aliryhmä, jos ah = Ha kaikilla a G. Toisin
Sovellettu todennäköisyyslaskenta B
 Sovellettu todennäköisyyslaskenta B Antti Rasila 8. marraskuuta 2007 Antti Rasila () TodB 8. marraskuuta 2007 1 / 18 1 Kertausta: momenttimenetelmä ja suurimman uskottavuuden menetelmä 2 Tilastollinen
Sovellettu todennäköisyyslaskenta B Antti Rasila 8. marraskuuta 2007 Antti Rasila () TodB 8. marraskuuta 2007 1 / 18 1 Kertausta: momenttimenetelmä ja suurimman uskottavuuden menetelmä 2 Tilastollinen
Harjoitus 7: NCSS - Tilastollinen analyysi
 Harjoitus 7: NCSS - Tilastollinen analyysi Mat-2.2107 Sovelletun matematiikan tietokonetyöt Syksy 2006 Mat-2.2107 Sovelletun matematiikan tietokonetyöt 1 Harjoituksen aiheita Tilastollinen testaus Testaukseen
Harjoitus 7: NCSS - Tilastollinen analyysi Mat-2.2107 Sovelletun matematiikan tietokonetyöt Syksy 2006 Mat-2.2107 Sovelletun matematiikan tietokonetyöt 1 Harjoituksen aiheita Tilastollinen testaus Testaukseen
Scolopendra morsitans -juoksujalkainen laji vai lajikompleksi? Johanna Hopkins
 Scolopendra morsitans -juoksujalkainen laji vai lajikompleksi? Johanna Hopkins Pro gradu -tutkielma Turun yliopisto Biologian laitos 27.3.2017 Linja: fysiologian ja genetiikan linja Erikoistumisala: evoluutiobiologia
Scolopendra morsitans -juoksujalkainen laji vai lajikompleksi? Johanna Hopkins Pro gradu -tutkielma Turun yliopisto Biologian laitos 27.3.2017 Linja: fysiologian ja genetiikan linja Erikoistumisala: evoluutiobiologia
LUOMINEN JA EVOLUUTIO
 LUOMINEN JA EVOLUUTIO Maailman syntyminen on uskon asia Evoluutioteoria Luominen Teoria, ei totuus Lähtökohta: selittää miten elollinen maailma olisi voinut syntyä, jos mitään yliluonnollista ei ole Ei
LUOMINEN JA EVOLUUTIO Maailman syntyminen on uskon asia Evoluutioteoria Luominen Teoria, ei totuus Lähtökohta: selittää miten elollinen maailma olisi voinut syntyä, jos mitään yliluonnollista ei ole Ei
2 vielä yleisimmin käytetty on 67 markkerin tutkimus, jolla testattavien välille löytyy yleensä jo eroja.
 DNA Ihmisessä on 30 biljoonaa solua. Solussa on useita osia, mm. tuma, jonka tehtävänä on säilyttää perinnöllistä aineistoa. Tuma sisältää kromosomipareja, joita ihmisellä on 23 kpl. Naisilla kaikki kromosomiparit
DNA Ihmisessä on 30 biljoonaa solua. Solussa on useita osia, mm. tuma, jonka tehtävänä on säilyttää perinnöllistä aineistoa. Tuma sisältää kromosomipareja, joita ihmisellä on 23 kpl. Naisilla kaikki kromosomiparit
Yhtäläisyydet selkärankaisten aivoissa, osa II. Niko Lankinen
 Yhtäläisyydet selkärankaisten aivoissa, osa II Niko Lankinen Sisältö Neuroneille tyypilliset molekyylit Suoraa jatkoa Niinan esitykseen Alkion aivojen vertailua Neuromeerinen malli Neuromeerisen mallin
Yhtäläisyydet selkärankaisten aivoissa, osa II Niko Lankinen Sisältö Neuroneille tyypilliset molekyylit Suoraa jatkoa Niinan esitykseen Alkion aivojen vertailua Neuromeerinen malli Neuromeerisen mallin
Aineistoista. Laadulliset menetelmät: miksi tarpeen? Haastattelut, fokusryhmät, havainnointi, historiantutkimus, miksei videointikin
 Aineistoista 11.2.09 IK Laadulliset menetelmät: miksi tarpeen? Haastattelut, fokusryhmät, havainnointi, historiantutkimus, miksei videointikin Muotoilussa kehittyneet menetelmät, lähinnä luotaimet Havainnointi:
Aineistoista 11.2.09 IK Laadulliset menetelmät: miksi tarpeen? Haastattelut, fokusryhmät, havainnointi, historiantutkimus, miksei videointikin Muotoilussa kehittyneet menetelmät, lähinnä luotaimet Havainnointi:
Etsintä verkosta (Searching from the Web) T Datasta tietoon Jouni Seppänen
 T Datasta tietoon Jouni Seppänen") Etsintä verkosta (Searching from the Web) T-61.2010 Datasta tietoon Jouni Seppänen 13.12.2006 1 Webin lyhyt historia 2 http://info.cern.ch/proposal.html 3 4 5 http://browser.arachne.cz/screen/ 6 7 Etsintä
Etsintä verkosta (Searching from the Web) T-61.2010 Datasta tietoon Jouni Seppänen 13.12.2006 1 Webin lyhyt historia 2 http://info.cern.ch/proposal.html 3 4 5 http://browser.arachne.cz/screen/ 6 7 Etsintä
Satunnaisalgoritmit. Topi Paavilainen. Laskennan teorian opintopiiri HELSINGIN YLIOPISTO Tietojenkäsittelytieteen laitos
 Satunnaisalgoritmit Topi Paavilainen Laskennan teorian opintopiiri HELSINGIN YLIOPISTO Tietojenkäsittelytieteen laitos Helsinki, 23. helmikuuta 2014 1 Johdanto Satunnaisalgoritmit ovat algoritmeja, joiden
Satunnaisalgoritmit Topi Paavilainen Laskennan teorian opintopiiri HELSINGIN YLIOPISTO Tietojenkäsittelytieteen laitos Helsinki, 23. helmikuuta 2014 1 Johdanto Satunnaisalgoritmit ovat algoritmeja, joiden