EPH-RESEPTORIN KLOONAUS, TUOTTO JA PUHDISTUS HYÖNTEISSOLUSTA
|
|
|
- Anton Hämäläinen
- 9 vuotta sitten
- Katselukertoja:
Transkriptio
1 Eph- reseptorin kloonaus, tuotto ja puhdistus hyönteissoluista Bio- ja Elintarviketekniikka Biotekniikka 2012 Shevin Mamandi EPH-RESEPTORIN KLOONAUS, TUOTTO JA PUHDISTUS HYÖNTEISSOLUSTA

2 OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio- ja elintarviketekniikka Biotekniikka Ilari Suominen, Sari Paavilainen Shevin Mamandi EPH-RESEPTORIN KLOONAUS, TUOTTO JA PUHDISTUS HYÖNTEISSOLUISTA Tämä opinnäytetyö tehtiin Joint Biotechnology Laboratoryssa osana Eph-reseptoreiden ja efriiniligandien sitoutusmis- ja rakennetutkimusta. Opinnäytetyössä EphA2-reseptoriproteiinin koko geeni kloonattiin pma152-a hyönteissoluvektoriin ja transformoitiin E.coli-soluhin. Opinnäytetyössä tuotettiin yhdistelmä- DNA-plasmidi yhdistelmä-dna-tekniikoilla. Työssä käytettyjä hyönteissoluja olivat Sf9 ja Hi5. Sf9 soluja käytettiin viruksen tuotossa ja Hi5 soluja käytettiin efriinia5-ligandiproteiinin sekä EphA2-reseptoriproteiinin tuotossa. Tuotettu efriini A5-ligandi puhdistettiin proteiini A-pylväällä ja EphA2-reseptori puhdistettiin Nipylväällä. Alkuvaiheessa EphA2 geeniin c-terminaaliin oltiin kiinnitetty histidiinihäntä, joka mahdollisiti proteiinin puhdistuksen nikkeligeelipylväällä. Analysointiin käytettiin yhdistelmä-dna-menetelmien aikana agaroosigeelielektroforeesia. Proteiinin tuotto vaiheessa näytteet analysointiin SDS-PAGE ja Western blotting menetelmillä. Aikaisemmissa opinnäytetöissä oli tuotettu ainostaan osaa Eph-reseptorista, mutta tässä opinnäytetyössä tuotettiin Eph-reseptorin koko geenin proteiinia ja analyysien perusteella proteiinin tuottaminen onnistui. ASIASANAT: (Eph-reseptorit, Hyönteissolut, Hyönteissoluvektori, Efriiniligandit)

3 BACHELOR S THESIS ABSTRACT TURKU UNIVERSITY OF APPLIED SCIENCES Biotechnology and Food Technology Biotechnology Ilari Suominen, Sari Paavilainen Shevin Mamandi EPH RECEPTOR CLONING, PRODUCTION AND PURIFICATION FROM INSECT CELLS This thesis was conducted at the Joint Biotechnology Laboratory as part of a research project concerning the attachment and conformation of Eph receptors and ephrin ligands. In this thesis, the whole gene coding for the EphA2-receptor protein was cloned into a pma152- a insect cell vector and transformed into E.coli cells. A recombinant DNA plasmid was produced by recombinant DNA plasmid methods. The insect cells used in this study were sf9 and Hi5. Sf9 cells were used for the production of a virus and Hi5 cells for the production of the EphA2 receptor and ephrin ligand proteins. The produced Ephrin-A5 ligand was purified with a protein A-column and the EphA2 receptor by affinity chromatography. A histidine tail was added to the C-terminal region of the EphA5 gene, which enabled the use of nickel affinity gel in the purification. Agarose gel electrophoresis was used for the analysis of recombinant DNA methods. SDS- PAGE and western blotting were used for the protein analysis. In previous Bachelor theses, parts of the Eph receptor have been produced. In the present study the entire gene coding for the protein was produced. According to the results the production was successful. KEYWORDS: (Insect cell, insect vectors, Eph-receptor, Ephrin ligand)

4 SISÄLTÖ 1 JOHDANTO Eph-reseptorit ja efriiniligandit Hyönteissolut Hyönteissoluvektori 9 2 MATERIAALIT JA MENETELMÄT PCR koko geenistä Digestio Ligaatio ja transformaatio Yhdistelmä-DNA-plasmidin analysointi digestiolla Hyönteissolujen kasvatus Bakulovirus Expressio systeemi P0-virus P1- virus P2-virus P3-virus EphA2:n tuotto Efriini-ligandin tuotto Lyysaus ja uutto Alkukoe Tuotoksen puhdistus affiniteettikromatografilla 27

5 2.10 Analysointi Agaroosigeelielektroforeesi SDS-page Western blottaus 28 3 TULOKSET JA TARKASTELU Koko geenin monistaminen PCR:llä Proteiinien tuotto ja puhdistus 33 4 YHTEENVETO 39 LÄHTEET 40 5 LIITTEET 43 KUVAT Kuva 1. EphA2-reseptorin kolmiulotteinen mallinnuskuva 3 8 Kuva 2. PacGP67-B bakulovirus transfektiovektorin geenikartta 8 10 Kuva 3. Yhdistelmä-DNA-plasmidin monistettava alue 19 Kuva 4. EphA2- geenin PCR 30 Kuva Minipreparaattien PCR 31 Kuva 6. Digestio 1,2,6 ja 10 Minipreparaateille 32 Kuva 7. Efriini- A5 proteiinin puhdistus 33 Kuva 8. Efriini A5 proteiinin puhdistus 33 Kuva 9. EphA2- reseptorin tuotto 34 Kuva 10. Alkukoe EphA2-proteiini tuotosta 35 Kuva 11. EphA2-reseptorin puhdistuksen affiniteettikromatogrammi 36 Kuva 12. EphA2-reseptorin puhdistus 37 Kuva 13.Western Blottaus EphA2:lle 37 TAULUKOT Taulukko 2. PCR putkien sisältö 12 Taulukko 3. PCR-ajo-ohjelma 13 Taulukko 4. Digestio pipetoinnit 15 Taulukko 5. Uutos laimennokset 26

6 KÄYTETYT LYHENTEET kda kilodalton EphA2-reseptor Ephrin type-a-reseptori 2 rpm PCR SDS-PAGE Sf9 Hi5 bp kierrosta minuutissa (revolutions per minute) polymeraasiketjureaktio (polymerase chain reaction) natriumlauryylisulfaattipolyakryyliamidigeelielektroforeesi hyönteissolukanta hyönteissolukanta emäspari

7 7 1 JOHDANTO Tämän opinnäytetyön tavoitteena oli kloonata, puhdistaa ja tuottaa EphA2- reseptoria Joint Biotechnolog Laboratoryssa rakennetutkimusta varten. Opinäytetyössä muodostettiin yhdistelmä-dna-tekniikoilla yhdistelmä-dnaplasmidi EphA2-geenistä ja hyönteissoluvektori pma152-a:sta. EphA2- reseptoria ja efriini A5-ligandia tuotettiin BEVS-menetelmällä käyttäen sf9 ja Hi5 hyönteissolulinjoja sekä tuotettua reseptoria ja ligandia puhdistettiin affiniteettikromatografialla. Analysointiin käytettiin agaroosigeelielektroforeesia, SDS-PAGE:a ja Western blottausta. 1.1 Eph-reseptorit ja efriiniligandit Tyrosiinikinaasireseptorit kuuluvat solukalvoproteiineihin ja tyrosiinikinaasit säätelevät toisten proteiinien aktiivisuutta fosforyloimalla proteiinien tyrosiineja. 1,2 Tyrosiinikinaasireseptoreiden suurperheeseen kuuluvat myös Eph-reseptorit ja efriiniligandi. Eph-reseptorit ja efriiniligandit jaetaan kahteen ryhmään A:han ja B:hen.Jako tapahtuu, joko niiden solun ulkoisten sekvenssien perusteella tai niihin sitoutuvien ligandien perusteella. Uudet näköhermot syntyvät, kun Ephreseptorit ja niiden ligandit ohjaavat uusien aksonien liikettä verkkokalvolla. Tutkimukset ovat myös osoittaneet, että ne osallistuvat keskushermoston synapsien muodostumiseen. 2,3, 4 Eph-reseptorit ovat proteiineja, jotka ovat kiinnittyneet solunkalvoon, ja on huomattu, että niiden singnalointi hillitsee aktiinin dynamiikkaa. Singnaalinvälityksellä reseptoreilla on kyky aktivoida useita solun osia samanaikaisesti. 2 Tässä opinäytetyössä kloonattiin EphA2-reseptorin koko geeni ja kyseistä reseptoria sekä efriini-a5-ligandia tuotettiin Hi5-hyönteissoluissa.

8 8 Kuva 1. EphA2-reseptorin kolmiulotteinen mallinnuskuva 3 Kuvassa 1 nähdään tietokoneella tehdyn EphA2-reseptorin kolmiulotteinen rakenne. Solu- ja molekyylitasolla ei vielä täysin käsitetä Eph- ja efriinimolekyylien vuorovaikutusta. Kuitenkin on todettu, että Eph-efriini-interaktiot ottavat osaa syöpäkasvainten ja verisuonten kehittymisessä säätelemällä niiden kriittisiä vaiheita, joten niitä käytettään syöpää diagnosoitaessa ja niitä pidetään kohteina syövän hoidolle. Ne ovat mukana kudosten ja elinten varhaisessa kehityksessä, mutta ne säätelevät myös elinten toimintaa myöhemmissä vaiheissa. 2 Jos Eph-reseptoreiden ja efriinien toimintaan voitaisiin vaikuttaa, saataisiin aikaan merkittäviä lääketieteelisiä saavutuksia. Näitä reseptoreita voitaisiin käyttää terapeuttisessa mielessä kasvattamalla esimerkiksi uusien hermosolujen määrää. 2

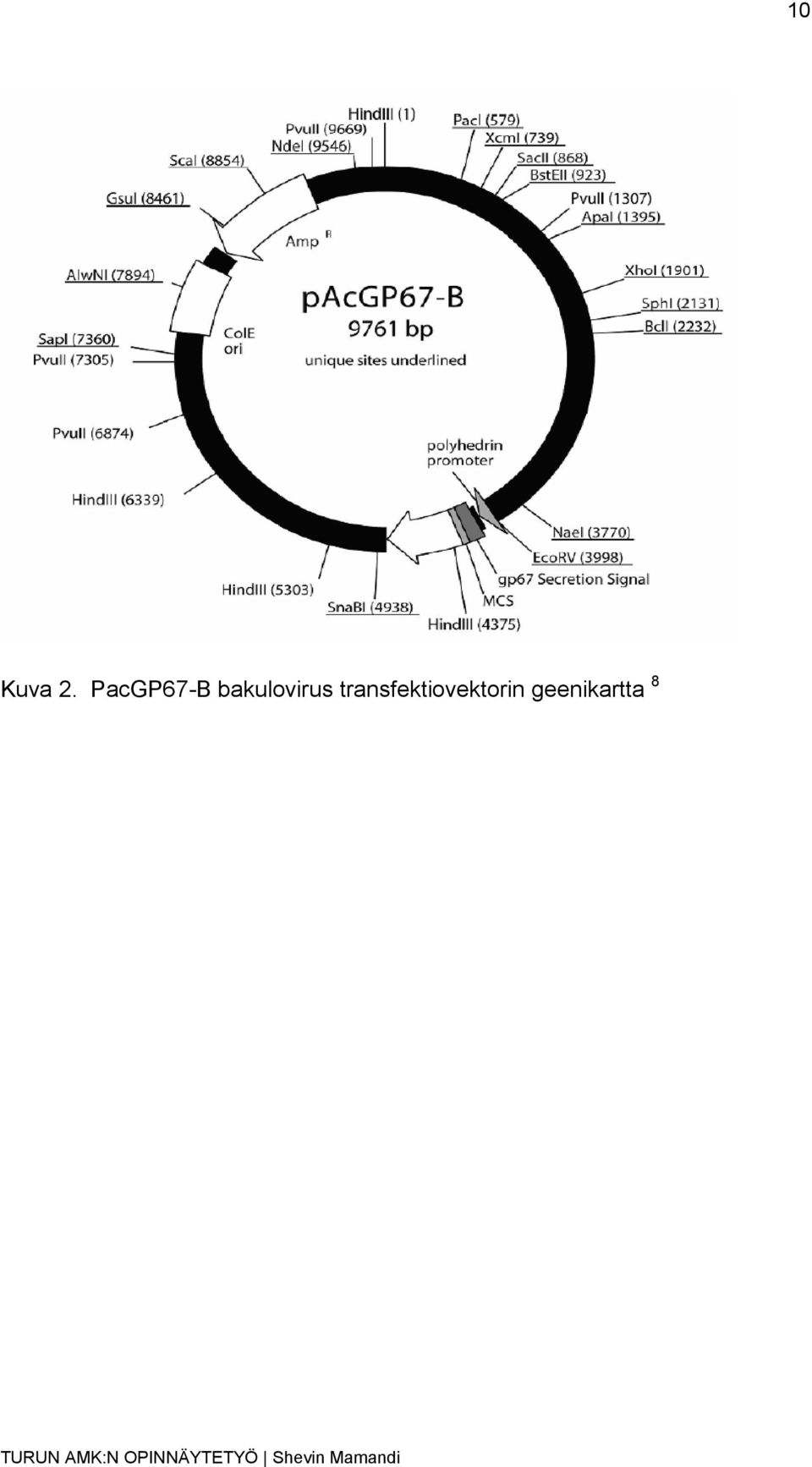
9 9 1.2 Hyönteissolut Hyönteissoluja käytetään humaaniproteiinin tuotannossa, koska niitä on helpompi viljellä ja ne ovat edullisempia kuin eläinsolut. Kun hyönteissoluun infektoidaan bakulovirus, se pystyy tuottamaan nopeasti suuria määriä haluttua proteiinia. Hyönteissoluilla ei ole insertin kokorajoitusta ja samanaikaisesti voidaan suoritta useampi geeniekspressio. 5,6 Opinnäytetyössä käyttämäni hyönteissolulinjat olivat Hi5-ja Sf9-soluja. Hi5 solut ovat hyönteisen munasarjan soluja ja Sf9-solut ovat sen kotelovaiheen munasarjan soluja. 5,6 1.3 Hyönteissoluvektori Hyönteissoluvektorit ovat sukkulavektoreita, joita monistetaan ja joiden yhdistelma-dna-plasmidit rakennetaan E. coli-bakteerisoluissa. Hyönteissoluvektorissa on ennen geenin alkua promoottori, joka mahdollistaa yhdistelmä-dna-plasmidin rekombinanttiproteiinin tuoton hyönteissoluissa. 10 (Sari Paavilainen, henkilökohtainen tiedonanto) Opinnäytetyössäni käytin bakulovirusvektoria, pma152-a, joka on muunneltu pacgp67-b:stä (kuva 2) ja on kooltaan 9761 kb. pacgp67-b hyönteissoluvektoriin on liitetty thrombiiniproteaasin katkaisukohdan lisäksi Myc- ja Fc-täki (engl. Fc-tag, Myc-tag), jolloin Fc-täkiä voidaan käyttää rekombinanttiproteiinin puhdistamisessa apuna. 10 (Sari Paavilainen, henkilökohtainen tiedonanto)

10 10 Kuva 2. PacGP67-B bakulovirus transfektiovektorin geenikartta 8
11 11 2 MATERIAALIT JA MENETELMÄT 2.1 PCR koko geenistä Polymeraasiketjureaktio (PCR eli polymerase chain reaction) on in vitromenetelmä, jolla voidaan kaksijuosteinen DNA-jakso monistaa eksponentiaalisesti. Menetelmä perustuu inkubaatioiden sarjaan, joka tehdään eri lämpötiloissa. Sillä voidaan monistaa pieni määrä DNA-jaksoa sopivaan konsentraatioon jatkotoimenpiteitä varten. 4,9,10 Kun PCR suoritetaan, tarvitaan siihen templaatti, jolla tarkoitetaan monistettavaa geenipätkää (tavallisesti kaksijousteinen DNA-pätkä). 10 PCR:n toimimiseen tarvitaan myös kaksi täysin tunnettua aluketta, jotka kiinnittyvät monistettavan geenipätkän alkuun ja loppuun eli siis koodattava geenipätkä jää alukkeiden sisälle. Alukkeet ovat kooltaan nukleotidin kokoisia fragmentteja, jotka käynnistävät kopioimisen. 10 DNA-polymeraasi kopioi templaatin sisältävän informaation muodostuvalle DNA-juosteelle. Polymeraasit ovat entsyymeja, jotka inaktivoituvat korkeissa lämpötiloissa, joten tämän takia PCR:ssä käytetään lämmönkestäviä polymeraaseja. DNA-polymeraaseja on kahdenlaisia, virheitä tekeviä ja vähemmän virheitä tekeviä eli oikolukuisia DNA-polymeraaseja. Oikolukuisia DNA-polymeraaseja suositaan enemmän PCR:ssä. 7,10 PCR koostuu kolmesta perusvaiheesta: denaturointi, annealing ja pidennysreaktio, jotka muodostavat yhdessä syklin. Denaturaatio suoritetaan viemällä lämpötila korkealle, jolloin DNA-juosteet irtoavat toisistaan. Annealing vaiheessa lämpötilaa lasketaan hieman, jolloin alukkeet kiinnittyvät DNAjuosteisiin. Tämän jälkeen suoritetaan pidennysreaktio nostamalla lämpötila noin 72 ⁰C:seen, jolloin putkessa olevat nukleotidit kiinnityvät templaatin kumpaankin juosteeseen DNA-polymeraasin ohjaamassa järjestyksessä. Kun molemmat nauhat on monistettu, niin nostetaan lämpötilaa, jolloin kaikki
.")
12 12 juosteet denaturoituvat noin 95⁰C:ssa ja seuraava sykli alkaa. Kun PCR suoritetaan jollekkin tuottelle, se koostuu yleensä syklistä riippuen siitä, kuinka paljon DNA:ta on käytettävissä ja paljonko sitä tarvitaan. 10 Työssä monistettiin PCR:n avulla EphA2-reseptoriproteiinin koko geeni, joka toimi templaattina PCR:ssä. Geeniin c-terminaaliin päähän liitettiin histidiinihäntä (histidiini-tag), jonka avulla pystyttiin helpommin puhdistamaan proteiini myöhäisemmässä vaiheissa. 10 Alukkeina käytettiin A2FL-BamH1-QGK-for (Oligomer oy, 18,0 nmol, 100,0 pmol/µl) ja A2/FL/His-tag-rev (Oligomer oy, 9,7 nmol, 100,0 pmol/µl). DNApolymeraasina käytettiin Phusion F-530s DNA-polymeraasia (Thermo, Lot 127, 2U/µl) ja nukleotideja 2mM dntp s (Pharmasia) sekä käytettiin 5x phusionpuskuriliuosta (Finnzymes, contains 7,5mM MgCl 2, Lot 66). Näytteitä valmistettiin PCR-reaktiota varten kahteen eri mikrosentrifuugiputkeen. Aineita pipetoitiin taulukon 1 mukaisesti, mutta toiseen putkista ei laitettu templaattia, koska se toimi kontrollina. Taulukko 1. PCR putkien sisältö Aine määrä (µl) H 2 O 24 2 mm dntp`s 5 EphA2-FC 1 F-530s DNA-polymeraasi 0,5 A2FL-BamHI-QGK-fo 5 A2/FL/His-tag 5 5x Phusion buffer 10
, jonka avulla pystyttiin helpommin puhdistamaan proteiini myöhäisemmässä vaiheissa.")
13 13 PCR ajoa varten tehtiin PCR-laitteelle oma ohjelma, joka on nähtävissä taulukossa 2. Taulukko 2. PCR-ajo-ohjelma Syklin vaihe Lämpötila Aika (min) (⁰C) Alku denaturointi 98 5 Denaturointi 98 1 Anniilaus 55 1 Ekstensio 72 3 Loppu ekstensio Lopetus 4 Syklien määrä: 35 Kun PCR-ajo oli päättynyt, tarkistettiin agaroosigeelielektroforeesilla, oliko tuotto onnistunut. Valmistettiin agaroosigeeli Liitteen 1 ohjeiden mukaan. Mikrosentrifuugiputkista otettiin 5 µl näytettä kahteen uuteen putkeen ja lisättiin niihin 2µl 6x DNA-LB-väriä, joka sisälsi bromifenolisinistä (väri) ja glyserolia. Glyseroli saa näytteet pysymään kaivossa tekemällä ne raskaammiksi. Agaroosielektroforeesigeelin kuoppiin pipetoitiin näytteet sekä niiden lisäksi kahteen kuoppaan pipetoitiin 5 µl λ-dna-standardia, joka oli digestoitu ECOR - ja Hind -restriktioentsyymeillä. λ-dna-standardi on markkeri, joka on digestoitu ja sen DNA-fragmenttien koot tiedetään. Ajo suoritettiin 100 voltilla noin 30 minuuttin ajan. Ajon jälkeen tarkistettiin, oliko PCR tuotteessa oikeata geeniä, joten geeli kuvaattiin (Kuva 4 ) geelinkuvantamislaitteella. 7,10 (Sari Paavilainen, henkilökohtainen tiedonanto) Haluttu geeni puhdistettiin kaupallisella spin-kolonilla Gene jet TM PCR purification kitillä (Fermentas, Lot ), johon on sidottuna kiinni
 ja glyserolia.")
14 14 silikamatriksi. Kaotroopin läsnäollessa silikamatrikisiin kiinnittyy nukleiinihapot suuressa ionivahvuudessa. 10 Ensin haluttu DNA-pala leikattiin agaroosigeelistä ja punnittiin. Geelipala sulatettiin lämpöblokissa ja sen jälkeen lisättiin sitoutumispuskuria (Fermentas, Lot ) yhtä paljon (eli suhteessa 1:1). Näytettä sentrifugoitiin noin 60 sekunttia, jolloin DNA tarttui silikaan. Kolonnista pestiin epäpuhtaudet 700 µl:lla pesupuskuria, johon oli ennen käyttöä lisätty 96 prosenttista etanolia ( , EVE), Fermentas, Lot ). Pesupuskurin lisäyksen jälkeen näytettä sentrifugoitiin noin 60 sekunttia. Pesu suoritettiin kahdesti. Pesun jälkeen geeni eluoitiin kolonnista 50 mikrolitralla eluointipuskuria (Fermentas, Lot ) ja sentrifugoitiin lopuksi 1minuutin ajan Digestio Digestiolla tarkoitetaan restriktioentsyymin käyttöä DNA-jakson katkaisemista varten. Restriktioentsyymillä voidaan katkaista insertti ja käytettävä plasmidi tietystä kohtaan (katkaisukohta on oltava käytettävän plasmidin rakenteessa), jolloin voidaan liittää insertti vektoriin ligaation avulla. Syy, miksi restriktioentsyymejä käytettiin yhdistelmä-dna-plasmidia muodostaessa, on se, että kyseiset entsyymit tunnistavat kaksinauhaisesta DNA:sta oikean nukleotidijärjestyksen, josta ne katkaisevat DNA:n. 10,12 Työssä restriktioentsyyminä käytettiin BamH :tä (Bacillus amyloliquefaciensbakteerikannan H restriktioentsyymi ) 10, jonka tunnistussekvenssit ovat: 5 GGATCC CCTAGG Tunnistuskohta rakennettiin inserttiin PCR:n avulla ja vektorina käytettiin pma152-a hyönteissoluvektoria, jossa oli valmiina tälle restiktioentsyymille katkaisukohta. 10
, Fermentas, Lot 00039252). Pesupuskurin lisäyksen jälkeen näytettä sentrifugoitiin noin 60 sekunttia. Pesu suoritettiin kahdesti.")
15 15 Työ suoritettiin pipetoimalla toiseen eppendorf-putkeen vektori, BamH puskuria (Fermentas, Lot ), Milli-Q-vettä ja restriktioentsyymiä BamH (Fermentas, Lot ) ja toiseen sama määrä tuotteita paitsi vektorin tilalle inserttiä. Tuotteita pipetoitiin eppendorf-putkiin taulukon 3 mukaisesti. Pipetoinnin jälkeen tuotteet sentrifugoitiin pohjaan ja näytteitä inkuboitiin lämpökaapissa 37 ⁰C:ssa kahden tunnin ajan. Taulukko 3. Digestio pipetoinnit Aine Määrä (µl) BamH -puskuri 3 BamH restriktioentsyymi 0,5 Milli-Q-vettä 16,5 Vektori tai Insertti 10 kokonaistilavuus 30 Inkuboinnin jälkeen restriktioentsyymi inaktivoitiin laittamalla putket 65 ⁰C:n lämpökuoppaan 15 minuutin ajaksi. Valmistettiin agaroosigeeli Liitteen 1 ohjeen mukaisesti. Ennen ajoa suoritettiin vektorille alkaalinen fosfataasi käsittely, jolla poistettiin vektorin päästä fosfaatti eli 5 -fosfaattiryhmä. Tämän toimenpiteen avulla ligaasin toiminta estetään ja vektorin päät eivät kiinnity ligaation aikana toistensa kanssa. Vektorin joukkoon pipetoitiin 1µl CIAP:ta (Calf Intestin Alkaline Phosphatase, Fermentas, #EF 0341, 1 U/µl, 200 U, lot 2524) ja inkuboitiin 35⁰C:ssa 30 minuutin ajan. Inkuboinnin jälkeen suoritettiin putkelle 85⁰ C:ssa lämpökäsittely lämpöblokissa 5 minuutin ajan, jolloin alkaalinen fosfataasi inaktivoituu. Insertin ja vektorin joukkoon laitettiin 6X DNA-LB-väriä ja ne pipetoitiin geelin kuoppiin. Yhteen kuoppaan pipetoitiin myös 5 µl λ-dna-standardia ja
 BamH -puskuri 3 BamH restriktioentsyymi 0,5 Milli-Q-vettä 16,5 Vektori tai Insertti 10 kokonaistilavuus 30 Inkuboinnin jälkeen restriktioentsyymi inaktivoitiin")
16 16 suoritettiin ajo 100 voltissa 30 minuutin ajan. Tämän jälkeen geeli kuvattiin, jotta nähtäisiin, onko digestio onnistunut. 2.3 Ligaatio ja transformaatio Ligaatiolla tarkoitetaan kahden DNA-jaksojen liittämistä toisiinsa entsyymien avulla kovalenttisin sidoksin. DNA-jaksojen kiinnittymisen mahdollistaa se, että niiden toisessa päässä on 5 -fosfaattiryhmä ja toisessa 3 -OH-ryhmä, jotka tunnistavat sopivat päät ja kiinnityyvät toisiinsa. Näitä ligaatiossa käytettyjä entsyymejä sanotaa DNA-ligaaseiksi. 10 Työssä käytettiin T4 DNA-ligaasia, joka on entsyymi, jota E.colin T4-faagi tuottaa. 10 Ligaatio suoritettiin pipetoimalla eppendorf-putkeen 2 µl digestoitua plasmidia, 6,5 µl digestoitua inserttiä (insertin ja vektorin suhteen on oltava noin 1:3 ), 1 µl 10x ligaatio puskuria (T4-buffer, Fermentas) sekä 0,5 µl T4 DNA ligaasia (Fermentas, #EL00II, Lot ). Kokonaistilavuus oli siis 10 µl. Yksi negatiivinen kontrolliputki tehtiin, jonne laitettiin kaikki samat kuin edellisessä putkessa, mutta insertin tilalle pipetoitiin Milli-Q-vettä. Näytteet sentrifugoitiin putkien pohjaan ja niitä inkuboitiin yksi tunti huoneenlämpötilassa. Tämän jälkeen ligaasi inaktivoitiin lämpöshokilla lämpöblokissa 65 ⁰C:ssa 15 minuutin ajan. Transformaatiolla tarkoitetaan geenin vaihtamista tai vieraan geenin tuottamista soluissa, jotka pystyvät ottamaan vastaan vierasta DNA:ta. Näitä soluja kutsutaan kompotenttisoluiksi ja ne on käsitelty kylmällä CaCl 2 :lla, jonka jälkeen niistä on tullut kompotentteja. 10 Transformaatio kompetentteihin E. coli-soluihin (XL1-blue, , EVE) aloitettiin ottamalla -70 C pakkasesta kaksi tilavuudeltaan noin 50 µl:n erää E. coli-soluja ja laittamalla ne suoraan jäihin, joista toiseen pipetoitiin 10 µl näytettä kontrolliputkesta ja toiseen putkeen pipetoitiin sama määrä putkesta, jossa on vektori ja siihen liitetty insertti. Putkia inkuboitiin jäissä noin 30 minuuttia, jonka jälkeen niille suoritettiin lämpöshokki 42 ⁰C:ssa 45 sekunnin ajan. Välittömästi

17 17 lämpöshokin jälkeen soluja inkuboitiin jäissä kahden minuutin ajan. Soluihin lisättiin inkubaation jälkeen SOC-mediumia, joka sisälsi 5,5 g tryptonia (Biokard diagnostics, Lot 9E524), 1,25 g hiivauutetta (Biokard diagnostics, Lot 3E946), 2,5 g NaCl (J.T.Baker, Lot ), 250 ml vettä ja 12,5 μg/ml tetrasykliiniä. Mediumin lisäyksen jälkeen soluja inkuboitiin ravistelijassa yhden tunnin ajan, jonka lämpötila oli 37 ⁰C ja ravistelunopeus oli 225 rpm. Tämän jälkeen suoritettiin molemmille putkille 10 0 ja 10-1 laimennokset, jotka pipetoitiin LAmaljoille. LA-maljoja varten valmistettiin agar, johon laitettiin 3,5 g Tryptonia (Biokard diagnostics, Lot 9E524), 1,25 g hiivauutetta (Biokard diagnostics, Lot 3E946), 2,5 g NaCl (J.T.Baker, Lot ), 250 ml vettä ja 3,75 g agaria (Becton Dickinson, Lot GM0084). Agar-liuos steriloitiin (20min, 121 C), ja jäähdytyksen jälkeen noin 50 C asteeseen, siihen lisättiin 1 ml ampisilliiniä (25mg/ml), jolloin LA-maljat valmistettiin kaatamalla agaria maljoille. Maljoja inkuboitiin yön yli 37⁰C:ssa. LA-maljoja käytettiin transformaatiossa sen takia, että vain ne solut kasvavat maljoilla, joissa on tapahtunut transformaatio eli ne sisältävät haluttua DNA:ta kiinnittyneenä plasmidiin. Muiden solujen, joissa ei ole plasmidia, kasvun estää ampisilliini, mutta solut, joissa on vektoria kasvavat, koska plasmidi tuottaa β-laktamaasia, joka tekee siitä ampisilliiniresistenssin. Kontrollilla testattiin, kuinka paljon transformaatiossa oli taustaa. Koska kontrollimaljalla oli vähemmän kasvua kuin toisessa maljassa (plasmidi + insertti), voitiin todeta, että transformaatio oli onnistunut. Minipreppien tekemisellä tarkoitetaan DNA:n eristämistä bakteeriviljelmästä. Minipreppien tekeminen aloitettiin transformaatiossa viljellyistä pesäkkeistä pipetoimalla ensin kymmeneen falconputkeen 5 millilitraa LB-mediumia (trypton 2.5g,hiivauute 1.25g, NaCl 2.5 g ja H 2 O 250ml ) ja 20µl ampisilliinia (25mg/ml). Tämän jälkeen otettiin maljalta (vektoria+inserttiä sisältältävät solut) 10 eri pesäkettä silmukalla ja liuotettiin ne falkkonputkiin. Pesäkkeitä inkuboitiin falkkonputkissa lämpöravistelijassa 37 ⁰C:ssa 250 rpm:ssä yön yli.

18 18 Falkkonputket sentrifugoitiin 4 ⁰C:ssa 4000 g:ssä 10 minuutin ajan. Tämän jälkeen supernatantit kaadettiin pois ja aloitettiin puhdistus GenElute TM plasmid miniprep-kitillä (Sigma, Lot PLN350), jolla erotettiin plasmidit soluista ja muista jätteistä. Pelletit liuotettiin 200 µl:an resuspensiopuskuria ja pipetoitiin liuotetut näytteet eppendorf-putkiin. Joukkoon lisättiin 200 µl lysis-puskuria (Sigma, Lot 037K6123) ja näytteitä sekoitettiin käsin kääntelemällä 3 minuuttia. Laitettiin joukkoon 350 µl sitoutumispuskuria (Sigma, Lot 047K6084) ja sentrifugoitiin 10 minuuttia g:ssä. Seuraavaksi otettiin 10 kollonnia, niihin pipetoitiin 500 µl preparation-puskuria (Sigma, Lot EA) ja niitä sentrifugoitiin yhden minuutin ajan g:ssä. Kolonneihin pipetoitiin näytteet ja niitä sentrifugoitiin yksi minuutti g:ssä. Supernatantit kaadettiin pois ja näytteet pestiin pipetoimalla jokaiseen kolonniin 750µl pesupuskuria (Sigma, Lot 037K6156). Näytteitä sentrifugoitiin yhden minuutin ajan g:ssä ja kaadettiin supernatantit pois. Pesu toistettiin kahdesti. Lopuksi plasmidit eluoitiin kolonneista eluointipuskurilla (Sigma, Lot 077K6172). Kolonneihin pipetoitiin 100µl eluointipuskuria, jonka jälkeen niitä sentrifugoitiin minuutin ajan g:ssä. 2.4 Yhdistelmä-DNA-plasmidin analysointi digestiolla Minipreparaateille suoritettiin PCR aiemmin käytetyn ohjelman mukaan ( Taulukko 2), jotta saataisiin selville, oliko eristettyjen minipreparaatien plasmideissa oikea geeni (EphA2). PCR:llä monistettiin 1500 bp:n mittainen A2ECD-geenipätkä EphA2-geenistä. Koe suoritettiin laittamalla reaktioputkeen alukkeet, joista toinen kiinnittyi EphA2-geenin ulkopuolelle vektoriin ennen kloonauskohtaa ja toinen kiinnittyi geenin sisäpuolelle (Kuva 3). PCR:n jälkeen valmistettiin agaroosigeeli Liitteen 1 ohjeiden mukaan ja ajo suoritettiin 100 voltissa noin 30 minuuttia. Ajon jälkeen geeli kuvattiin (Kuva 5 ).
 ja näytteitä sekoitettiin käsin kääntelemällä 3 minuuttia.")
19 19 Kuva 3. Yhdistelmä-DNA-plasmidin monistettava alue Minipreparaatit digestoitiin restriktioentsyymillä BamH. Digestion tarkoitus oli testata, oliko minipreparaateissa oikeaa inserttiä. Kun yhdistelmä-dnaplasmidit digestoitiin samalla restriktioentsyymillä kuin aikaisemmin, saatiin niistä oikean kokoinen DNA fragmentti näkyviin, kun agaroosielektroforeesi geeli oli kuvattu. (Sari Paavilainen, henkilökohtainen tiedonanto) Työ aloitettiin pipetoimalla 7µl ydistelmä-dna-plasmidia, 2 µl BamH1 puskuria (Fermentas, Lot ), 10.5 µl milli-q-vettä ja 0,4 µl restriktioentsyymiä BamH ( Fermentas, Lot ) eppendorf-putkiin. Digestion jälkeen näytteisiin lisättiin 4 µl 6xDNA-LB-väriä ja pipetoitiin 12µl näytettä kuoppiin. Yhteen kuoppaan pipetoitiin myös 5 µl digestoitua λ-dna:ta kontrolliksi. Valmistettiin geeli Liitteen 1 ohjeiden mukaisesti. Ajettiin geeliä noin 30 minuuttia, jonka jälkeen se kuvattiin (Kuva 6) ja todettiin, että osassa näytteistä näytti olevan oikeata inserttiä. Tämän jälkeen lähetettiin neljä yhdistelmä-dna-
 eppendorf-putkiin. Digestion jälkeen näytteisiin lisättiin 4 µl 6xDNA-LB-väriä ja pipetoitiin 12µl näytettä kuoppiin.")
20 20 plasmidia sekvensoitavaksi biotekniikan keskuksen sekvensointipalveluun (Biocityn 5 kerrokseen) Hyönteissolujen kasvatus Otettiin Hi5- ja sf9-hyönteissoluampullit nestetypestä ja herätettiin ne henkiin seuraavalla tavalla. Sulatettiin käsissä 1 ml:n Hi5-hyönteissoluampulli, pipetoitiin se 250 ml erlenmeyeriin, johon lisättiin 20ml Insect Xpress-mediumia ja lopuksi inkubointiin elenmeyeriä 27⁰C:ssa 100rpm:ssä muutaman päivän ajan. Hi5 solut kaksinkertaistuvat noin vuorokaudessa. (Sari Paavilainen, henkilökohtainen tiedonanto) Sf9-hyönteissoluampulli sulatettiin (STA,NY ) samoin kuin Hi5-solut, mutta ampullin sisältö pipetoitiin petrimaljalle ja solujen päälle lisättiin 5 ml Insect SF-4 Baculo Express kasvuliuosta (promcell, Lot E05196P). Maljaa inkuboitiin yhden tunnin ajan 27 ⁰C:ssa, jotta solut tarttuvat maljan pohjalle. Yhden tunnin inkubointi johtuu siitä, että solut oli pakastettu kasvuliuokseen, joka sisältää DMSO:ta. Se säilyttää solut pakasteessa hyvin, mutta sulaneena se on haitallinen soluille. Tämän takia soluliuos pipetoitiin maljalta uudelle maljalle ja solujen päälle lisättiin uutta mediumia. Toiselle maljalle suoritettin sama tunnin inkubointi, joka jälkeen pipetoitiin soluliuos pois ja laitettiin uutta mediumia tarttuneiden solujen päälle. Sf9-solut kaksinkertaistuivat noin kahdessa vuorokaudessa. (Sari Paavilainen, henkilökohtainen tiedonanto) Solujen tiheyttä pidettiin 0,5-6,0 miljoonaa solua/ml ja ne laimennettiin joka toinen päivä 0,8 miljoonaa solua/ml, jolloin ne pärjäävät muutaman päivän. Solujen kasvatuksen aikana laimennettaessa niihin lisättiin myös penisilliini/streptomysiini (Pen/ strep, penicilliini streplomycin, Lot Be12-730Q) antibioottia. (Sari Paavilainen, henkilökohtainen tiedonanto)
 Sf9-hyönteissoluampulli sulatettiin (STA,NY 29.8.")
21 21 Solut laskettiin pipetoimalla Bürker-kammioon 10 µl soluliuosta, jolloin saadaan laskettua mikroskoopilla kammion ruudukoiden avulla, kuinka paljon soluja on 1 millilitrassa soluliuosta. 2.5 Bakulovirus Expressio systeemi Hyönteissoluja voidaan infektoida bakuloviruksella, joka ei lisäänny hyönteissolun ulkopuolella. Ne ovat rakenteeltaan kapselimaisia ja ne sisältävät sirkulaarisen genomin. BEVS-menetelmässä (Bakulovirus Expressio systeemi) vektorina on bakulovirus, jota tuotetaan hyönteissoluissa ja siirretään toiseen hyönteissoluun tuotettavaksi. Bakuloviruksen avulla pystytään tuottamaan runsaasti haluttua proteiinia lyhyessä ajassa. 6,13 Haluttua bakulovirusta tuotettiin neljässä vaiheessa, jonka jälkeen se siirrettiin hyönteissoluun, jossa haluttua proteiinia (EphA2) tuotettiin. Kun virukset pipetoidaan solujen joukkoon, tunkeutuvat virukset endosytoosin avulla aktiivisiin soluihin ja infektoidessaan, soluissa alkaa monistumaan virusta kunnes solujen tumat turpoavat ja solujen jakautuminen päättyy. Tämä johtaa solujen kuolemaan. 6,14 Opinnäytetyössä tuotettiin virus ja se infektoitiin hyönteissoluun eli suoritettiin BEVS eli bakulovirusekspressio. Näistä kaikista vaiheista otettiin näytteitä ja lopuksi suoritettiin SDS-PAGE määritys Liitteen 2 ohjeiden mukaisesti P0-virus Pipetoitiin kuoppalevylle (Cell star greinen bio-one lot E , Cat , 24 well cell culture plate) sf9-hyönteissoluja kahdeksaan kuoppaan (3.31 miljoonaa solua/ml) 0.67 miljoonaa solua per kuoppa eli 202µl ja laitettiin joukkoon sf-4-baculo express mediumia (with L-glutamine) lot E0519p).Inkuboitiin 27 ⁰C:ssa yhden tunnin ajan.
22 22 Inkuboinnin aikana otettiin kaksi eppendorfputkea, joista toiseen putkeen pipetoitiin 3µl plasmidia, jossa oli insertti. Samaan putkeen pipetoitiin myös 1µl BaculoGoldia (linearized baculovirus DNA, Lot 53226) ja inkuboitiin viisi minuuttia. Inkuboinnin jälkeen lisättiin 40µl grace s insect mediumia (unsupplemented 1x, GIBCO 11595, Lot ). Toiseen tuubiin pipetoitiin 40µl grace s insect mediumia ja 4.2µl cellfectin reagenssia (inritrogen, Lot ). Molempien eppendorf-putkien sisältö yhdistettiin ja inkuboitiin huoneenlämpötilassa 30 minuutin ajan. Kuoppalevyltä pipetoitiin (aikaisemmin oli pipetoitu kahdeksaan kuoppaan sf9-soluja ja sf-4-express-mediumia ja inkuboitu niitä yksi tunti) medium pois ja pestiin kuopat 400 µl:lla grace s miediumia. Medium pipetoitiin pois ja ykkös- sekä kakkoskuoppiin pipetoitiin 204 µl/kuoppa eppendorfputken (sisältää yhdistelmä-dna-plasmidin sekä BaculoGold-DNA:n eli näytteen) sisällöstä. Loppuihin kuuteen kuoppaan pipetoitiin 204µl grace s mediumia, joita käytettiin kontrollina. Näytteitä inkuboitiin 27 ⁰C:ssa viiden tunnin ajan ja vaihdettiin kuoppien medium 400 µl:an sf4-mediumia (promcell, Lot E05196P). Viruksen kehittymistä tarkkailtiin päivittäin. Kun infektoituneet solut alkoivat vähitellen turvota ja kuolla, poistettiin infektoituneet solut ensimmäisestä ja toisesta kuopasta kahteen falkkonputkeen ja sentrifugoitiin niitä viiden minuutin ajan 4000 rpm:ssä. Supernatantit säilytettiin 4 ⁰C:ssa P1- virus. Otettiin kuuden kuopan kuoppalevy ja pipetoitiin jokaiseen kuoppaan 3 miljoonaa sf9-soluja (1.64 miljoonaa solua/ ml) eli 1.9 ml ja inkuboitiin yhden tunnin ajan. Pipetoitiin kuoppiin yksi ja kaksi 450 µl P0-virusta, 2,55 ml sf4- mediumia sekä 30µl penicilliini streptomyciniä. Kontrolleiksi otettiin loput neljä kuoppaa, joihin pipetoitiin vain 3 ml sf4-mediuimia sekä 30 µl penicilliini streptomyciniä. Inkuboitiin kuoppalevy 27 ⁰C:ssa niin kauan, että infektoituneet solut alkoivat vähitellen turvota ja kuolla. Tämän jälkeen poistettiin infektoituneet solut ensimmäisestä ja toisesta kuopasta kahteen falkkonputkeen ja
23 23 sentrifugoitiin viiden minuutin ajan 4000 rpm:ssä. supernatantit säilytettiin 4 ⁰C:ssa P2-virus Otettiin 15cm halkaisijaltaan oleva malja ja pipetoitiin siihen 36 miljonaa sf9- solua ja inkuboitiin maljaa yhden tunnin ajan, jotta solut tarttuisivat maljan pohjaan. Inkuboinnin jälkeen maljalle pipetoitiin 1 ml P1-virusta ja 25 ml Insect express mediumia sekä 250 µl penicilliini streptomyciniä. Maljaa inkuboitiin 27⁰C:ssa, kunnes soluissa tapahtui muutos niinkuin aikaisemmissa virustuotoissa (P0- ja P1-virus), jonka jälkeen raaputettiin maljalta solut irti ja pipetoitiin ne falkkonputkiin ja sentrifugoitiin viiden minuutin ajan 4000 rpm:ssä. Osa supernatantista otettiin talteen ja pakastetiin P3-virus Pipetoitiin isolle maljalle (cellsfar, tissue culture ps, 145x20 mm, Lot EIQ1008x,Cat ) 40 miljoonaa solua ja inkuboitiin tunnin ajan, jotta solut tarttuisivat pohjaan. Kun solut olivat tarttuneet maljan pohjaan, pipetoitiin tarttumattomat solut pois. Pipetoitiin maljalle 1 ml P2-virusta, 20ml insect express mediumia sekä 250 µl penicilliini streptomyciniä. Inkuboitiin maljaa 27⁰C:ssa, kunnes soluissa tapahtui muutos, jonka jälkeen solut raaputettiin maljalta irti. Pipetoitiin ne falkkonputkiin ja sentrifugoitiin viiden minuutin ajan 4000 rpm:ssä, jonka jälkeen supernatantti otettiin talteen proteiinin tuottoa varten EphA2:n tuotto Pipetoitiin 6ml P3-virusta Hi5-solujen (300 ml) joukkoon ja laitettiin kasvamaan 100rpm:n 27⁰C:n. Kun solut olivat kasvaneet muutaman päivän, solut
24 24 sentrifugoitiin 4500 rpm:ssä 4⁰C:ssa 30 minuutin ajan. Supernatantti otettiin talteen ja laitettiin jäihin odottamaan. 2.6 Efriini-ligandin tuotto Hi5 hyönteissolussa tuotettiin efriiniligandia (Turun yliopistossa biologiaa opiskeleva tyttö, joka työskenteli JBL:ssä oli suorittanut kaikki geenin kloonauksen työvaiheet, joita on tehty tässä opinäytetyössä EphA2-reseptorin geenille), joka aktivoi EphA2-proteiinia. 2 Otettiin erlenmeyer-pullo, johon laitettiin 400 ml Hi5-soluja ja 400ml Insect-Xpress mediumia sekä 8 ml penicilliini streptomycini-antibioottia. Erlenmeyerpullon sisältö jaettiin kahteen erlenmeyerpulloon, lisättiin kumpaankin 8 ml virusta (Efrin A5 p4, , Laura) ja inkuboitiin lämpöravistelijassa 48 tuntia 27 ⁰C:ssa 100 rpm:ssä. Molemmista erlenmeyerpulloista otettiin 1ml:n näyte analysointia varten (sentrifugoidaan ennen analysoimista 10 minuuttia 4000rpm:ssä). Inkuboinnin jälkeen pullot sentrifugoitiin 4000 g:ssä 30 minuutin ajan ja supernatantti otettiin talteen. Sentrifugointi suoritettiin, jotta saataisiin efriini-a5-ligandiproteiini erotettua Hi5 soluista. Efriini puhdistettiin proteiini A-agaroosipylväällä (rproteine A sepharose gel ), johon tarttuu efriinissä oleva Fc-täki (engl. Fc-tag). (Sari Paavilainen, henkilökohtainen tiedonanto) Tarvittavat liuokset olivat valmiina sekä pylväs oli koottu ja tasapainoitettu valmiiksi. Proteiini A-pylväässä oleva 20% etanoli pestiin Milli-Q-vedellä virtausnopeudella 1ml/minuutti ja tasapainoitettiin 10ml aloituspuskuria, PROT A-puskuria (20mM Hepes, 0,5 mm NaCl, 2,5 mm MgCl 2, ph 7.3, Sari). Tasapainotuksen jälkeen ajettiin 730 ml:n efriini-kasvatus virtausnopeudella 0,5 ml/minuuttia 22 tunnin ajan pylvään läpi. Ajon jälkeen otettiin talteen läpitullut fraktio siltä varalta, että efriini ei olisi tarttunut pylvääseen. Pylväs pestiin 10 ml:lla tasapainoituspuskuria. Huuhtominen suoritettiin, koska tasapainoituspuskuri sisälsi NaCl:a, joka estää
25 25 ionivarauksiin perustuvan tarttumisen pylvääseen eli epäspesifisen tarttumisen. Näyte eluoitiin 10ml:lla eluointipuskuria (0,1 M glysiini, 0.5 M NaCl, 2,5 mm MgCl 2, ph3, ,Sari) ja fraktiot kerättiin kymmeneen eppendorfputkeen (2ml fraktio/putki). 2.7 Lyysaus ja uutto Lyysauksella tarkoitetaan solujen hajoittamista palasiksi, jolloin proteiini on helpompi poistaa soluosista uuttamalla. 11 EphA2-reseptorin tuottovaiheen solut pestiin 30 ml:lla pesupuskuria (10mM Hepes, 20mM KCl, 10mM MgCl 2 ) (medumia ja muuta pestiin pois) ja sentrifugoitiin rpm:ssä 30 minuutin ajan. Tämän jälkeen liuotettiin solut 20 ml:an lysis-puskuria (50mM Hepes, 1M NaCl, 1M KCl) solut imevät sisälleen puskuria ja lisättiin joukkoon 50 µl proteaasi-inhibiittoria (Sigma fast protease inhibitor, Lot ), joka estää proteaasia plkkomasta tuotettavaa proteiinia. Mukaan laitettiin myös 100µl DNAasia (100U/mg, , Sari), jota käytettiin, koska haluttiin sen pilkkovan DNA:ta pois. (Sari Paavilainen, henkilökohtainen tiedonanto) Solut sentrifugoitiin rpm:ssä 30 minuutin ajan, jonka jälkeen kaadettiin supernatantti (otettiin ainostaan analysoimista varten 1ml talteen) pois. Pellettiä uutettiin 20ml:lla lyysis-puskuria, johon lisättiin myös 100 µl proteaasi inhibiittoria. Joukkoon laitettiin myös 40mg kolesteryyli-vety-sukkinaatti (Mpbio, Lot 9519H) ja 100mg n-dodekyyli-β-d-maltosidia (Biotop, Lot Cu633). Ne ovat detergenttejä, jotka hajottavat solun membraanirakennetta ja proteiinit alkavat irrota solukalvosta. Pellettejä uutettiin sekoittajassa yön yli ja seuraavana päivänä ne sentrifugoitiin 55 minuutin ajan rpm:ssä. Otettiin supernatantista talteen 1ml analysointia varten. Suorititettiin uutto prosessi vielä kertaalleen eli näytteitä uutettiin kahdesti.
26 Alkukoe Nikkeligeelikoe tehtiin uutoksille, koska sillä varmistuttiin, oliko niissä tuotettua proteiinia. Siitä voitiin myös päätellä, että kiunka paljon sitä oli näytteessä. (Sari Paavilainen, henkilökohtainen tiedonanto) Pipetoitiin eppendorf-putkeen 300 µl nikkeligeeliä (Midazde, mm,sisältää 20 % etanolia, Lot ), koska se sisälsi etanolia (haittaa proteiinin tartuumista nikkeligeeliin). Geeli pestiin kahdesti lisäämällä siihen 800 µl:a milli- Q-vettä ja se sentrifugoitiin 6000 rpm:ssä kahden minuutin ajan. Otettiin näytteet uutoksista ja laimennettiin ne taulukon 7 mukaisesti. Pipetoinnin jälkeen eppendorf-putkia inkuboitiin sekoittajassa 45 minuutin ajan. Taulukko 4. Uutos laimennokset pipetoitiin Uutto 1 Uutto 1 Uutto 2 Uutto 2 (U1.1) (U1.2) (U2.1) (U2.2) (µl) (µl) (µl) (µl) Uutos IMAC Nikkeli geeli Inkuboinnin päätyttyä sentrifugoitiin eppendorf-putket 6000 rpm:ssä kahden minuutin ajan ja supernatantti kaadettiin pois. Liuotettiin pelletti 500 µl:an IMAC 30 (20mM hepes, 0,5mM KCl, 30mM imidatsoli, 2,5mM MgCl 2, ,EVA) puskuria ja sentrifugoidaan 6000 rpm:ssä kahden minuutin ajan. Kahden liuotuksen jälkeen kaadettiin supernatantti pois ja liuotettiin putkissa oleva pelletti 10µ:lla SDS-Page väriä. Putket laitettiin lämpöblokkiin 10 minuutiksi 60 ⁰C:seen ( lämpöblokilla denaturoidaan näytteet).
27 27 Liiteen 2 ohjeen mukaan valmistettiin ylägeeli(kasausgeeli) ja alageeli(erotusgeeli) sekä suoritettiin SDS-Page ajo. 2.9 Tuotoksen puhdistus affiniteettikromatografilla Affiniteettikromatografia perustuu proteiinin kinnittymiseen matriksiin heikoin spesiifisin sidoksin. Tässä opinnäytetyössäni käytin Ni-pylvästä, jossa on postitiivisesti varautuneita Ni +2 -ioneja. Nämä ioinit sitovat EphA2-reseptorin monistusvaiheessa kiinnitetyn histidiinihännän. IMAC-puskurin laimella imidatsolipitoisuudella saadaan reseptorin histidiinihäntä kiinnittymään ja korkealla imidatsolipitoisuudella irtoamaan matriksista. 12,18 Tuotos puhdistetaan 1 ml:n nikkelipylväällä (Ni sepharose TM 6 fast flow contains 20% ethanol presevrative, GE Healthcare, Lot ). Pylväs puhdistettiin 20%:lla etanolilla virtausnopeudalla 0,5 ml/min 30 minuutin ajan. Tämän jälkeen huuhdeltiin pylväs milli-q-vedellä samalla virtausnopeudella yhden tunnin ajan. Pylväs tasapainoitettiin IMAC-30 puskuriilla (20mM hepes, 0,5M KCl, 30 mm imidatsoli, 2.5mM MgCl 2, 0.05% DDM ja 0,01% CHS, Sari, ). Ajettiin näyte virtausnopeudella 0,5 ml/min ja eluoitiin tarttumaton proteiini ajon jälkeen IMAC-30 puskurilla (sama kuin tasapainoituksessa) nopeudella 1 ml/min, kunnes kaikki tarttumaton proteiini oli tullut ulos (seurattiin piirturilla, kunnes perustaso oli nolla). Kun tarttumaton proteiini saatin ulos, eluoitiin EphA2-his proteiini ulos IMAC-500 puskurilla (20mM Hepes,pH 7,3, 0,5M KCl, 2.5 mm MgCl 2, 500mM imidasoli, 0,05 % DDM, 0,01% CHS, Sari, ). Lopuksi puhdistettin pylväs vedellä ja 20 %:lla etanolilla Analysointi Agaroosigeelielektroforeesi Elektroforeesi perustuu sähkövirtaan, joka erottelee molekyylit toisistaan. Agroosigeelin avulla DNA-pätkät kulkeutuvat sähkövirrassa.
28 28 Agaroosielektroforeesissa on plus- ja miinusnapa. DNA sisältää fosfaattiryhmiä, jotka saavat DNA:n varautumaan negatiivisesti ja sen kulkeutumaan kohti miinusnapaa sähkövirrassa. Agaroosin geeliydyttyä, se muodostaa verkkorakenteen, jossa pienemmät fragmentit kulkevat nopeammin ja isommat puolestaan liikkuvat hitaammin, jolloin fragmentien pituus saadaan selville kuvattaessa geeliä. Geeliin valmistusvaiheessa siihen lisätään etidiumbromidia, jolla se värjäytyy. Kun tutkitaan geeliä UV-valossa ja kuvaataan se kuvantamislaitteella, fragmentit nähdään oranssina, joka johtuu siitä, että etidiumionit ovat tunkeutuneet DNA:ssa nukleiinihappojen emästen väliin. 4, SDS-page SDS-PAGE on menetelmä, jonka avulla analysoidaan ja erotetaan proteiineja toisistaan. 4,15,18 SDS-PAGE:n (natriumdodekyylisulfaattipolyakryyliamidigeelielektroforeesi) toiminnan perustana on agaroorielektroforeesin tapaan sähkövirta. Agaroosi geelin tilalla SDS-PAGE:ssa käytettään polyakryyiamiidigeeliä, jossa huokoskoko on samaa luokkaa proteiinien kanssa. Siksi tämä menetelmä on hyvä tapa erottaa haluttu proteiini muista proteiineista. Tämä menetelmä perustuu agaroosielektroforeesin tapaan molekyylikoon perusteella erotteluun. SDS-PAGE:ssa SDS on detergentti, joka on negatiivisesti varautunut ja se saa proteiinit nauhamaiseksi muuttamalla ne negatiivisesti varautuneiksi. 4,15,18 PAGE:n geeli koostuu ylägeelisitä (kasausgeeli) ja alageelistä (erotusgeeli). Geeli valmistettiin ja SDS-PAGE ajo suoritettiin Liitteen 1 ohjeen mukaan Western blottaus Western blot on menetelmä, jolla voidaan tunnistaa proteiini, joka on erotettu SDS-PAGE avulla. Erotettu proteiinin vyöhyke kinnitettään vasta-aineen avulla membraaniin. 17,18
29 29 Leikattiin neljä sopivankokoista Whatman-paperia ja leikattiin yksi membraani (Bio-Rad, immun-blot TM PVDF membrane 0,2µl, Lot BR ), jolla proteiini blotattiin. Membraani aktivoitiin laittamalla se viideksi sekunniksi metanoliin, jonka jälkeen se laitettiin veteen. Reikäteline koottiin laittamalla sen väliin yksi geeli, kaksi sientä, neljä watman paperia sekä yksi membraani. Ajopuskuria (1x Trasfer Buffer) kaadettiin ajoastiaan. Geeli ajatettiin 100 voltissa yhden tunnin ajan, jonka jälkeen se laitettiin maitoon (1xPBS, 5% maitojauhetta, KCl, NaCl,KH 2 PO 4, Na 2 HPO 4 ) yön yli. Seuraavaksi pestiin membraani 1xPBS puskurilla ( , Laura) ja laitettiin ravistelijaan viideksi minuutiksi. Tämän jälkeen membraania inkuboitiin yhden tunnin ajan penta-his vasta-ainetta sisältävässä liuoksessa, jolloin se tunnistaa proteiinin C-terminaaliin kiinnitetyn histidiinitäkin. (Sari Paavilainen, henkilökohtainen tiedonanto) Inkuboinnin jälkeen pestiin kolmesti membraani 1xPDS + Tween puskurilla ( , EPÄ), joka saa proteiinin hohtamaan pimeässä. Kehitettiin filmi 500µl:lla ECl-liuoksella [Detection reagent 2 luminol enhancer solution (prod num PIERCE, Lot ) ja detection reagent 1 (peroxide solution, prod. num , PIERCE, Lot )] ja 500µl:lla western blotting substraatilla.
30 30 3 TULOKSET JA TARKASTELU 3.1 Koko geenin monistaminen PCR:llä Kuva 4. EphA2- geenin PCR Kuvassa 4 on kuvattu agaroosigeelielektroforeesi PCR-ajon tulokset. Std tarkoittaa λ-dna-standardia, joka on digestoitu ja sen fragmenttien koot tiedetään (listattu fragmenttien koko kuvan oikealla puolella). Ins tarkoitta puolestaan inserttiä eli EphA2-reseptorin geeniä. EphA2-geeni on noin 2900 bp:ä ja kuvasta voidaan päätellä, että insertin vyöhyke on kooltaan lähes 2900 bp:ä, kun verrataan sitä standardin vyöhykkeisiin. Kuvan perusteella voidaan todeta, että monistaminen PCR:llä on onnistunut.
31 31 Kuva Minipreparaattien PCR Kuvassa 5 on kuvattu kymmenen minipreparaatin PCR-reaktioiden agaroosigeelielektroforeesi. Standardina oli digestoitu λ-dna. PCR suoritettiin, jotta saataisiin selville, oliko minipreparaateissa oikeaa geeniä oikeinpäin. A2ECD-geeni, jota PCR:ssä monistettiin, oli kooltaan noin 1500 pb:tä. Kuvan perusteella havaitaan kyseinen vyöhyke näytteiden 1,2 ja 10 kohdalla. Minipreparaatit, joissa ei näy vyöhykettä, niissä ei ole EphA2-geeniä ollenkan tai jos geeni on, niin se ei ole oikeinpäin.
32 32 Kuva 6. Digestio 1,2,6 ja 10 Minipreparaateille Minipreparaatit, joiden kuvan 2 perusteella todettiin sisältävän kyseistä geeniä sekä minipreparaatti 6, jossa ei näkynyt geeniä, digestoitiin BamH1 restriktioentsyymillä. Digestion jälkeen suoritettiin agaroosigeelielektroforeesiajo, joka kuvattiin. Kuvassa nähdään, että kaikissa miniprepeissä näkyy vyöhyke, joten voidaan todeta, että kaikissa mukaan lukien 6, on insertti, mutta 6:ssa insertti on väärinpäin.
33 Proteiinien tuotto ja puhdistus Kuva 7. Efriini- A5 proteiinin puhdistus Kuva 8. Efriini A5 proteiinin puhdistus Hi5 hyönteissoluissa tuotettu efriini-a5-ligandi puhdistettiin proteiini A- agaroosipylväällä ja siitä ajettiin SDS-PAGE, jotta saatiin selville, missä vaiheessa puhdistusta puhdistettava proteiini tuli ulos. Ennen puhdistusta
34 34 Efriiniligandia oli tuotettu Hi5 hyöntessolussa kahdessa erlenmeyer-pullosta, joista kummastakin otettiin näytteet (1-2) ja (1-4). Ft on supernatantti, joka on tullut pylvään läpi kiinnittymisvaiheessa. Näytteille Ft (Flow- through), (1-2) ja (1-4) suoritettiin puhdistus proteiinia A-agaroosilla. Näytteiden joukkoon laitettiin 40 µl proteiini A-agaroosia ja niitä inkuboitiin huoneen lämpötilassa yhden tunnin ajan. Kuvissa 7 ja 8 näytteet 1-13, joista vyöhyke on muodostunut näytteisiin 3-12 eli proteiinia on tullut ulos näihin eppendorf-putkiin. Kuvan perusteella voidaan päätellä, että eniten proteiinia oli tullut ulos 5-8 näytteistä. (1-2) ja (1-4) näytteissä näyttäisi olevan proteiinia ja myös Ft näytteessä näkyy vyöhyke, joka näyttäisi olevan samalla tasolla oikean proteiinin kanssa, joten kaikki efriinia5 ei kiinnittynyt pylvääseen. Kuva 9. EphA2- reseptorin tuotto Kuvassa 9 on kuvattu EphA2-proteiinin tuotto eri vaiheissa. Sf9- ja Hi5- hyönteissolut otettiin kontrolleiksi ja kuvasta huomattiin, että niissä oli paljon proteiinia, mutta oikean proteiinin kohdalla ei ole vyöhyhettä, joten niissä ei ole
35 35 EphA2-proteiinia. N (P3 virus + Hi5-solut ) on puolestaan EphA2-reseptorin tuottovaiheesta Hi5-hyönteissolussa ja siinä näyttää olevan paljon muuta proteiinia kuten myös EphA2-proteiinia (noin 120 kda). P3 on näyte P3- viruksen tuottovaiheesta ja siitä huomataan, että siinä näkyy kunnon vyöhyke oikean proteiinin kohdalla (120 kda). Kuva 10. Alkukoe EphA2-proteiini tuotosta SDS-PAGE määritys suoritettiin EphA2-reseptorin alkukoe- ja tuottovaiheista (katso Kuva 10). Hi5 solut, jotka sisälsivät EphA2 proteiinia, uutettiin kahdesti, joista otettiin näytteet (U1 ja U2) uuttojen välillä. Uutoksille tehtiin vielä laimennokset (U1.1, U1.2,U2.1 ja U2.2 ) katso Taulukko 3. Kuvasta nähdään, että kaikissa uutos-näytteissä oli proteiinin vyöhyke, joka on 120 kda:n kohdalla. Uutoksista kuitenkin laimeammassa on vaalemapi vyöhyke sekä uutos 1:ssä että 2:ssa. SN sisältää pesupuskuria, Hi5 soluja sekä EphA2- proteiinia ja siinä näyttäisi olevan vyöhyke 120 kda:n kohdalla, mutta joukossa on paljon muutakin proteiinia. SNT sisältää samaa kuin SN paitsi, että näyte oli myös lyysattu. Siinä näyttäisi olevan proteiinia runsaasti mukaanlukien haluttua (EphA2) proteiinia. N puolestaan sisälsi Hi5-hyönteissoluja ja EphA2-reseptoria. N:ssä proteiinivyöhykkeitä oli runsaasti, jonka takia halutun proteiinin vyöhykettä ei pysty erottamaan muista.

36 36 U1 U2 Kuva 11. EphA2-reseptorin puhdistuksen affiniteettikromatogrammi Kuvassa 11 on kuvattu affiniteettikoromatografilla puhdistettu EphA2-reseptori. Uutetut proteiinit puhdistettiin kahdessa erillisessä ajossa, joiden välissä pylväs puhdistettiin ja tasapainoitettiin. Tasapainoituksen ja puhdistuksen aikana piirturi pysäytettiin. Ensin ajettiin kerran uutettu näyte (U1) ja sen jälkeen ajettiin kaksi kertaa uutettu (U2). Kromatogrammissa nähdään kaksi piikkiä, joista voidaan todeta, että ensimmäisen uuton proteiini on tullut ulos ensimmäisen piikin (U1) kohdalla ja toinen piiki kuvaa toisen uutoksen (U2) puhdistusta.
37 37 Kuva 12. EphA2-reseptorin puhdistus Kuva 13.Western Blottaus EphA2:lle Kuvassa 12 ajettiin SDS-PAGE selvittääksemme, oliko tuotettu proteiini puhdistunut. Huomattiin, että proteiinia oli seuraavissa näytteissä U1, U2, N1 ja N2, koska 120 kda kohdalla näkyi vyöhyke. Hi5-soluissa oli runsaasti muuta proteiinia. N:ssä näkyi myös runsaasti muuta proteiinia, mutta myös 120 kda:n kohdalla näkyi vyöhyke. Ft eli pylväästä ulos tulleessa supernatantissa näyttäisi
38 38 olevan todella vähän tai ei ollenkaan proteiinia, joten voidaan todeta, että puhdistus on onnistunut. Kuvassa 13 oli samat näytteet ajettu samassa puskurissa, mutta se blotattiin membraanin penta-his-vasta-aineella (histidiinivasta-aine). Huomattiin, että ylhäällä toteamamme tulokset olivat oikein eli N1, N2, U1 ja U2 sisältävät EphA2-reseptoria. Ft puolestaan ei sisältänyt EphA2:ta, joten voidaan sanoa, että puhdistus oli täysin onnistunut.
39 39 4 YHTEENVETO Opinnäytetyön tavoitteena oli kloonata, tuottaa ja puhdistaa EphA2-reseptoria hyönteissoluista. Koska EphA2-reseptorin geeni on kooltaan pitkä, niin se kloonattiin hyönteissoluista peräisin olevaan vektoriin, sillä hyönteissoluilla voi tuotta suurista DNA-pätkistä proteiinia. Yhdistelmä-DNA-tekniikoilla yhdistelmä-dna-plasmidin tuottaminen onnistui, mutta ligaatiossa sekä transformaatiossa oli jonkinlaisia ongelmia. Ongelma muodostui siitä, että vektori kiinnittyi itsensä kanssa ligaatiossa. Ongelma ratkaistiin suorittamalla vektorille alkaalinen fosfataasi-käsittely, jolloin sen fosfaattipää poistettiin ja sillä estettiin sen kiinnittymistä itsensä kanssa. Muuten kaikki työvaiheet toimivat tehokkaasti. Tavoitteena oli suorittaa tuotetuille EphA2-reseptoreille ja efriinia5-ligandille sitoutumiskokeita, mutta ajanpuutteen takia se työvaihe jätettiin pois. Opinnäytetyön käytännönosa oli pitkä prosessi ja kirjoittaminen oli haastavaa, mutta suuren kiitoksen ansaitsee ohjaajani Sari Paavilainen, joka jaksoi olla kärsivällinen ja auttoi minua kaikin voimin.
40 40 LÄHTEET 1. Solunetti (2006), Tyroksiinkikinaasireseptorit [online viitattu, ] Saatavilla: 2. Himanen J.-P. & Nikolov D. B. (2003) Saatavilla: ph%20receptors%20and%20ephrins.pdf, The international Journal of B iochemistry & Cell Biology vol.35 sivut Wikipedia,Kuva EphA2 [online viitattu ] Saatavilla: 4. Päivänsäde E.(2009) EphA3-reseptoriproteiinien ligandia sitovan osan tuotto eri isäntäorganismeissa sekä puhdistus ja karakterisointi. Opinnäytetyö. Turun Ammattikorkeakoulu, Bio-ja elintarviketekniikka. 5. Ikonomou L. & Schneider Y.-J.& Agathos S. N., 2003, Insect cell culture for industrial production of recombinant proteins, Appl Microbiol Biotechnol vol. 62 sivut 1-21 [viitattu, ] Saatavilla: xtpdf/ eb5b2/1?accountid= Susi Anastasia, 2011, Virusproteiinien tuotto hyönteissoluissa bakuloviruksen avulla.opinnäytetyö. Turun Ammattikorkeakoulu. Bio-ja elintarviketekniikka. 7. Kojo H.-L. & Kemppainen N. &Keränen P., 2009, PCR-menetelmän optimointi kihomadon DNA:n osoittamiseksi ulosteesta, Pirkanmaan Ammattikorkeakoulu [viitattu, ] Saatavilla: n.pdf?sequence=1
41 41 8. Kuva (vektori) [viitattu, ] Saatavilla: b_enabled/21221p_ pdf 9. Wikipedia, PCR, [online viitattu ] Saatavilla: Suominen I. & Pärssinen R. & Haajanen K. & Pelkonen J.(2010), Geenitekniikka, Saarjärven Offset Oy, Saarijärvi Vainiopää M. (2011)Mikrobien cdna:n synteesin automatisointi ribosomaalisesta RNA:sta, Opinnäytetyö Jyväskylän Ammattikorkeakoulu.Laboratorioala Saatavilla: marianna.pdf?sequence=1 12. Aitomäki E. & Eerikäinen T. & Leisola M. ym.(2002), Bioprosessitekniikka, WSOY, Helsinki 13. Wikipedia, Bakulovirus, [viitattu, ] Saatavilla: Virtanen Sanna Pauliina, 2008, β1,3-glukosyylitransferaasin tuoton optimointi ja puhdistus, Bakulovirukset [viitattu, ] Saatavilla: (Virtanen S. (2008), Progradu-tutkielma. Tampereen yliopisto. Lääketieteelisen teknologian instituutti.) 15. Solunetti, 2006, SDS-PAGE [online, viitattu, ], Saatavilla: Solunetti, 2006,Western Blottaus [online, viitattu, ], Saatavilla: Solunetti, 2006, Digestio [viitattu, ] Saatavilla:
Eeva Seppänen ENDOTELIINI-1-FUUSIOPROTEIININ KLOONAAMINEN JA TUOTTAMINEN
 Eeva Seppänen ENDOTELIINI-1-FUUSIOPROTEIININ KLOONAAMINEN JA TUOTTAMINEN ENDOTELIINI-1-FUUSIOPROTEIININ KLOONAAMINEN JA TUOTTAMINEN Eeva Seppänen Opinnäytetyö Syksy 2010 Laboratorioalan koulutusohjelma
Eeva Seppänen ENDOTELIINI-1-FUUSIOPROTEIININ KLOONAAMINEN JA TUOTTAMINEN ENDOTELIINI-1-FUUSIOPROTEIININ KLOONAAMINEN JA TUOTTAMINEN Eeva Seppänen Opinnäytetyö Syksy 2010 Laboratorioalan koulutusohjelma
PCR - tekniikka elintarvikeanalytiikassa
 PCR - tekniikka elintarvikeanalytiikassa Listerian, Salmonellan ja kampylobakteerien tunnistus elintarvikkeista ja rehuista 29.11.2012 Eva Fredriksson-Lidsle Listeria monocytogenes Salmonella (spp) Campylobacter
PCR - tekniikka elintarvikeanalytiikassa Listerian, Salmonellan ja kampylobakteerien tunnistus elintarvikkeista ja rehuista 29.11.2012 Eva Fredriksson-Lidsle Listeria monocytogenes Salmonella (spp) Campylobacter
Plasmidi-DNA:n eristys bakteerisoluista DNA:n geelielektroforeesi (Proteiinien geelielektroforeesi)
") Plasmidi-DNA:n eristys bakteerisoluista DNA:n geelielektroforeesi (Proteiinien geelielektroforeesi) CHEM-A1310 Biotieteen perusteet Heli Viskari 2017 DNA-harjoitustöiden aikataulu, valitse yksi näistä
Plasmidi-DNA:n eristys bakteerisoluista DNA:n geelielektroforeesi (Proteiinien geelielektroforeesi) CHEM-A1310 Biotieteen perusteet Heli Viskari 2017 DNA-harjoitustöiden aikataulu, valitse yksi näistä
GEENITEKNIIKAN PERUSASIOITA
 GEENITEKNIIKAN PERUSASIOITA GEENITEKNIIKKKA ON BIOTEKNIIKAN OSA-ALUE! Biotekniikka tutkii ja kehittää elävien solujen, solun osien, biokemiallisten menetelmien sekä molekyylibiologian uusimpien menetelmien
GEENITEKNIIKAN PERUSASIOITA GEENITEKNIIKKKA ON BIOTEKNIIKAN OSA-ALUE! Biotekniikka tutkii ja kehittää elävien solujen, solun osien, biokemiallisten menetelmien sekä molekyylibiologian uusimpien menetelmien
Genetiikan perusteiden harjoitustyöt
 Genetiikan perusteiden harjoitustyöt Molekyylien kloonaus ja siihen liittyvät taidot ja temput, osa 1 Restriktioentsyymit, elektroforeesi Moniste sivulta 24-: Geenien kloonaus CELL 491- Isolating, cloning,
Genetiikan perusteiden harjoitustyöt Molekyylien kloonaus ja siihen liittyvät taidot ja temput, osa 1 Restriktioentsyymit, elektroforeesi Moniste sivulta 24-: Geenien kloonaus CELL 491- Isolating, cloning,
LABORATORIOTYÖ: AGAROOSIGEELIELEKTROFOREESI
 LABORATORIOTYÖ: AGAROOSIGEELIELEKTROFOREESI Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNAjaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Eri valmistajien
LABORATORIOTYÖ: AGAROOSIGEELIELEKTROFOREESI Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNAjaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Eri valmistajien
Bioteknologian perustyökaluja
 Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
EPHA3-RESEPTORIPROTEII I LIGA DIA SITOVA OSA TUOTTO ERI ISÄ TÄORGA ISMEISSA SEKÄ PUHDISTUS JA KARAKTERISOI TI
 Opinnäytetyö EPHA3-RESEPTORIPROTEII I LIGA DIA SITOVA OSA TUOTTO ERI ISÄ TÄORGA ISMEISSA SEKÄ PUHDISTUS JA KARAKTERISOI TI Eerika Päivänsäde Bio- ja Elintarviketekniikka 2009 TURUN AMMATTIKORKEAKOULU TIIVISTELMÄ
Opinnäytetyö EPHA3-RESEPTORIPROTEII I LIGA DIA SITOVA OSA TUOTTO ERI ISÄ TÄORGA ISMEISSA SEKÄ PUHDISTUS JA KARAKTERISOI TI Eerika Päivänsäde Bio- ja Elintarviketekniikka 2009 TURUN AMMATTIKORKEAKOULU TIIVISTELMÄ
Tenttipäivät. Tentti TI :30-17:30 1. uusinta MA :30-17:30 2. uusinta MA :30-17:30
 Tenttipäivät Tentti TI 14.11. 14:30-17:30 1. uusinta MA 11.12. 14:30-17:30 2. uusinta MA 30.1.2018 14:30-17:30 Jos on joku iso päällekkäisyys (= tutkinnon pakollisen kurssin pakollinen läsnäolo, esimerkiksi
Tenttipäivät Tentti TI 14.11. 14:30-17:30 1. uusinta MA 11.12. 14:30-17:30 2. uusinta MA 30.1.2018 14:30-17:30 Jos on joku iso päällekkäisyys (= tutkinnon pakollisen kurssin pakollinen läsnäolo, esimerkiksi
10.9.2015 Pipetointi, sentrifugointi ja spektrofotometria
 BIOKEMIAN MENETELMÄT I, SYKSY 2015 VASTAUKSET LUENTOMATERIAALIN TEHTÄVIIN: 10.9.2015 Pipetointi, sentrifugointi ja spektrofotometria Pesukoneen g-voimat: RCF (x g) = 1,119 10-5 (rpm)2 r = roottorin säde
BIOKEMIAN MENETELMÄT I, SYKSY 2015 VASTAUKSET LUENTOMATERIAALIN TEHTÄVIIN: 10.9.2015 Pipetointi, sentrifugointi ja spektrofotometria Pesukoneen g-voimat: RCF (x g) = 1,119 10-5 (rpm)2 r = roottorin säde
Biokemian menetelmät I kurssi, työselostukset, kevät 2016.
 Biokemian menetelmät I kurssi, työselostukset, kevät 2016. DEADLINET: työselostus tulostettuna paperille Työ 3: To 24.3.2016 klo 15:00 KE1132:n palautuspiste tai BMTK:n Työ 2: Pe 1.4.2016 klo 16:00 KE1132:n
Biokemian menetelmät I kurssi, työselostukset, kevät 2016. DEADLINET: työselostus tulostettuna paperille Työ 3: To 24.3.2016 klo 15:00 KE1132:n palautuspiste tai BMTK:n Työ 2: Pe 1.4.2016 klo 16:00 KE1132:n
MYKOPLASMA- WORKSHOP!
 Science whereyou are Tarjoukset voimassa 31.12.2014 asti kampanjakoodilla FIN311214FIN. Kampanjahinnat ilman arvonlisäveroa. TULOSSA... MYKOPLASMA- WORKSHOP! BioNordika Finland Oy Kutomotie 18, 00380 Helsinki
Science whereyou are Tarjoukset voimassa 31.12.2014 asti kampanjakoodilla FIN311214FIN. Kampanjahinnat ilman arvonlisäveroa. TULOSSA... MYKOPLASMA- WORKSHOP! BioNordika Finland Oy Kutomotie 18, 00380 Helsinki
CHEM-A1310 Biotieteen perusteet 2018
 CHEM-A1310 Biotieteen perusteet 2018 Laboratoriodemojen teoriaosuus Huomaa: Jokaisesta suorittamatta jääneestä demokerrasta vähennetään 0.75 pistettä tentin kokonaispisteistä. Rästikertojen jälkeen suoritukset
CHEM-A1310 Biotieteen perusteet 2018 Laboratoriodemojen teoriaosuus Huomaa: Jokaisesta suorittamatta jääneestä demokerrasta vähennetään 0.75 pistettä tentin kokonaispisteistä. Rästikertojen jälkeen suoritukset
TITRAUKSET, KALIBROINNIT, SÄHKÖNJOHTAVUUS, HAPPOJEN JA EMÄSTEN TARKASTELU
 Oulun Seudun Ammattiopisto Raportti Page 1 of 6 Turkka Sunnari & Janika Pietilä 23.1.2016 TITRAUKSET, KALIBROINNIT, SÄHKÖNJOHTAVUUS, HAPPOJEN JA EMÄSTEN TARKASTELU PERIAATE/MENETELMÄ Työssä valmistetaan
Oulun Seudun Ammattiopisto Raportti Page 1 of 6 Turkka Sunnari & Janika Pietilä 23.1.2016 TITRAUKSET, KALIBROINNIT, SÄHKÖNJOHTAVUUS, HAPPOJEN JA EMÄSTEN TARKASTELU PERIAATE/MENETELMÄ Työssä valmistetaan
Juha Laitinen. Itsemurhavektorin valmistaminen Yersinia enterocolitica O:3:n ligaasigeenin inaktivoimista varten
 Juha Laitinen Itsemurhavektorin valmistaminen Yersinia enterocolitica O:3:n ligaasigeenin inaktivoimista varten Metropolia Ammattikorkeakoulu Laboratorioanalyytikko (AMK) Laboratorioala Opinnäytetyö 27.11.2011
Juha Laitinen Itsemurhavektorin valmistaminen Yersinia enterocolitica O:3:n ligaasigeenin inaktivoimista varten Metropolia Ammattikorkeakoulu Laboratorioanalyytikko (AMK) Laboratorioala Opinnäytetyö 27.11.2011
TRPA1-PROTEIININ TUOTTO JA KARAKTERISOINTI WESTERN BLOT -MENETELMÄLLÄ
 TRPA1-PROTEIININ TUOTTO JA KARAKTERISOINTI WESTERN BLOT -MENETELMÄLLÄ Meiju Kukkonen Kehittämistehtävä Kesäkuu 2010 Solu- ja molekyylibiologian erikoistumisopinnot Tampereen ammattikorkeakoulu TIIVISTELMÄ
TRPA1-PROTEIININ TUOTTO JA KARAKTERISOINTI WESTERN BLOT -MENETELMÄLLÄ Meiju Kukkonen Kehittämistehtävä Kesäkuu 2010 Solu- ja molekyylibiologian erikoistumisopinnot Tampereen ammattikorkeakoulu TIIVISTELMÄ
CHEM-A1310 Biotieteen perusteet 2019
 CHEM-A1310 Biotieteen perusteet 2019 Laboratoriodemojen teoriaosuus Huomaa: Jokaisesta suorittamatta jääneestä demokerrasta vähennetään 0.75 pistettä tentin kokonaispisteistä. Rästikertojen jälkeen suoritukset
CHEM-A1310 Biotieteen perusteet 2019 Laboratoriodemojen teoriaosuus Huomaa: Jokaisesta suorittamatta jääneestä demokerrasta vähennetään 0.75 pistettä tentin kokonaispisteistä. Rästikertojen jälkeen suoritukset
you Science Tarjoukset voimassa 30.6.2015 asti kampanjakoodilla kevät15. Kampanjahinnat ilman arvonlisäveroa. are
 Science where you are Tarjoukset voimassa 30.6.2015 asti kampanjakoodilla kevät15. Kampanjahinnat ilman arvonlisäveroa. BioNordika Finland Oy Kutomotie 18, 00380 Helsinki tel 0207 410 270 info@bionordika.fi
Science where you are Tarjoukset voimassa 30.6.2015 asti kampanjakoodilla kevät15. Kampanjahinnat ilman arvonlisäveroa. BioNordika Finland Oy Kutomotie 18, 00380 Helsinki tel 0207 410 270 info@bionordika.fi
Tuija Solismaa IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS
 Tuija Solismaa IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS RAPORTIN NIMIÖSIVU IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS
Tuija Solismaa IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS RAPORTIN NIMIÖSIVU IHMISEN RECQL4-PROTEIININ 450 ENSIMMÄISTÄ AMINOHAPPOA KOODITTAVAN DNA-JAKSON KLOONAUS
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
9/30/2013. GMO analytiikka. Termistöä. Markkinoilla olevien GM kasvien ominaisuuksia
 GMO analytiikka Kemian ja toksikologian tutkimusyksikkö Evira Termistöä geenimuuntelu muuntogeeninen siirtogeeninen GM GMO (geneettisesti muunnettu organismi) GM tapahtuma (event): käytetään silloin kun
GMO analytiikka Kemian ja toksikologian tutkimusyksikkö Evira Termistöä geenimuuntelu muuntogeeninen siirtogeeninen GM GMO (geneettisesti muunnettu organismi) GM tapahtuma (event): käytetään silloin kun
Biokemian labrameiningit I harjoitustyöosuus. Arne Raasakka, 20.10.2007 Työ suoritettu: 12. 13.10.2007 arne.raasakka@oulu.fi
 Työ 1. Rekömbinanttipröteiinin puhdistaminen Ni-NTA affiniteettikrömatögrafialla seka pröteiinin mölekyylipainön ma a ritys elektröföreesilla ja geelisuödatuskrömatögrafialla Biokemian labrameiningit I
Työ 1. Rekömbinanttipröteiinin puhdistaminen Ni-NTA affiniteettikrömatögrafialla seka pröteiinin mölekyylipainön ma a ritys elektröföreesilla ja geelisuödatuskrömatögrafialla Biokemian labrameiningit I
Geenitekniikan perusmenetelmät
 Loppukurssikoe To klo 14-16 2 osiota: monivalintatehtäväosio ja kirjallinen osio, jossa vastataan kahteen kysymykseen viidestä. Koe on auki klo 14.05-16. Voit tehdä sen oppitunnilla, jolloin saat tarvittaessa
Loppukurssikoe To klo 14-16 2 osiota: monivalintatehtäväosio ja kirjallinen osio, jossa vastataan kahteen kysymykseen viidestä. Koe on auki klo 14.05-16. Voit tehdä sen oppitunnilla, jolloin saat tarvittaessa
Torque teno viruksen proteiinien tuotto ja puhdistus
 Torque teno viruksen proteiinien tuotto ja puhdistus Bioanalytiikan koulutusohjelma, bioanalyytikko Opinnäytetyö 16.4.2007 Eija Lönn HELSINGIN AMMATTIKORKEAKOULU STADIA Sosiaali- ja terveysala Koulutusohjelma
Torque teno viruksen proteiinien tuotto ja puhdistus Bioanalytiikan koulutusohjelma, bioanalyytikko Opinnäytetyö 16.4.2007 Eija Lönn HELSINGIN AMMATTIKORKEAKOULU STADIA Sosiaali- ja terveysala Koulutusohjelma
TEKNIIKKA JA LIIKENNE. Laboratorioala
 TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ LACTOBACILLUS SOBRIUS -KANNAN PINTAKERROSPROTEIINIGEENIN KLOONAUS, ILMENTÄMINEN JA PROTEIININ KIINNITTYMISOMINAISUUDET Työn tekijä: Anne Fortelius Työn
TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ LACTOBACILLUS SOBRIUS -KANNAN PINTAKERROSPROTEIINIGEENIN KLOONAUS, ILMENTÄMINEN JA PROTEIININ KIINNITTYMISOMINAISUUDET Työn tekijä: Anne Fortelius Työn
LABORATORIOTYÖ: RESTRIKTIOENTSYYMIDIGESTIO
 LABORATORIOTYÖ: RESTRIKTIOENTSYYMIDIGESTIO Restriktioentsyymidigestio on yksi yhdistelmä-dna-tekniikan perusmenetelmistä, jossa katkaistaan kaksinauhainen DNA tietystä kohdasta erilaisten restriktioentsyymien
LABORATORIOTYÖ: RESTRIKTIOENTSYYMIDIGESTIO Restriktioentsyymidigestio on yksi yhdistelmä-dna-tekniikan perusmenetelmistä, jossa katkaistaan kaksinauhainen DNA tietystä kohdasta erilaisten restriktioentsyymien
VIRUSPROTEIINIEN TUOTTO HYÖNTEISSOLUISSA BAKULOVIRUKSEN AVULLA
 Opinnäytetyö (AMK) Bio- ja elintarviketekniikka Biotekniikka 2011 31 s Anastasia Susi VIRUSPROTEIINIEN TUOTTO HYÖNTEISSOLUISSA BAKULOVIRUKSEN AVULLA OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU
Opinnäytetyö (AMK) Bio- ja elintarviketekniikka Biotekniikka 2011 31 s Anastasia Susi VIRUSPROTEIINIEN TUOTTO HYÖNTEISSOLUISSA BAKULOVIRUKSEN AVULLA OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU
Pihlajanrengaslaikkuvirus Suomessa
 Pihlajanrengaslaikkuvirus Suomessa Ida Laine Maisterintutkielma Helsingin Yliopisto Maataloustieteiden laitos Kasvipatologia 2015 Tiedekunta/Osasto Fakultet/Sektion Facult Maatalous-metsätieteellinen tiedekunta
Pihlajanrengaslaikkuvirus Suomessa Ida Laine Maisterintutkielma Helsingin Yliopisto Maataloustieteiden laitos Kasvipatologia 2015 Tiedekunta/Osasto Fakultet/Sektion Facult Maatalous-metsätieteellinen tiedekunta
GMO analytiikka Annikki Welling Kemian tutkimusyksikkö Evira
 GMO analytiikka Annikki Welling Kemian tutkimusyksikkö Evira Millaisia GM kasvit ovat ja kuinka tätä käytetään hyväksi analytiikassa Aromaattisten aminohappojen biosynteesireitti kasvissa Kasvi tarvitsee
GMO analytiikka Annikki Welling Kemian tutkimusyksikkö Evira Millaisia GM kasvit ovat ja kuinka tätä käytetään hyväksi analytiikassa Aromaattisten aminohappojen biosynteesireitti kasvissa Kasvi tarvitsee
α-amylaasi α-amylaasin eristäminen syljestä ja spesifisen aktiivisuuden määritys. Johdanto Tärkkelys Oligosakkaridit Maltoosi + glukoosi
 n eristäminen syljestä ja spesifisen aktiivisuuden määritys. Johdanto Työssä eristetään ja puhdistetaan merkittävä ja laajalti käytetty teollisuusentsyymi syljestä. pilkkoo tärkkelystä ensin oligosakkarideiksi
n eristäminen syljestä ja spesifisen aktiivisuuden määritys. Johdanto Työssä eristetään ja puhdistetaan merkittävä ja laajalti käytetty teollisuusentsyymi syljestä. pilkkoo tärkkelystä ensin oligosakkarideiksi
Maija Joensuu & Maija Hanhela NAV3-GEENIMERKKIAINEMENETELMÄN KEHITTÄMISPROJEKTI
 Maija Joensuu & Maija Hanhela NAV3-GEENIMERKKIAINEMENETELMÄN KEHITTÄMISPROJEKTI NAV3-GEENIMERKKIAINEMENETELMÄN KEHITTÄMISPROJEKTI Maija Hanhela & Maija Joensuu Opinnäytetyö Kevät 2015 Bioanalytiikan koulutusohjelma
Maija Joensuu & Maija Hanhela NAV3-GEENIMERKKIAINEMENETELMÄN KEHITTÄMISPROJEKTI NAV3-GEENIMERKKIAINEMENETELMÄN KEHITTÄMISPROJEKTI Maija Hanhela & Maija Joensuu Opinnäytetyö Kevät 2015 Bioanalytiikan koulutusohjelma
COXSACKIEVIRUS A9 KANTOJEN MUUNTUMINEN
 Opinnäytetyö (AMK) Bio ja elintarviketekniikka Biotekniikka 2011 Tiina Kelanne COXSACKIEVIRUS A9 KANTOJEN MUUNTUMINEN VP1- ja 3D-geenien sekvensointi rekombinaatioiden tunnistamiseksi OPINNÄYTETYÖ (AMK)
Opinnäytetyö (AMK) Bio ja elintarviketekniikka Biotekniikka 2011 Tiina Kelanne COXSACKIEVIRUS A9 KANTOJEN MUUNTUMINEN VP1- ja 3D-geenien sekvensointi rekombinaatioiden tunnistamiseksi OPINNÄYTETYÖ (AMK)
SUBKLOONAUS JA RESTRIKTIOENTSYYMIANALYYSI
 SUBKLOONAUS JA RESTRIKTIOENTSYYMIANALYYSI Molekyylibiologian ja geeniteknologian harjoitustyön kokoaminen Savoniaammattikorkeakoulun bioanalytiikan koulutusohjelmalle Opinnäytetyö Maija Vuorenpää Bioanalytiikan
SUBKLOONAUS JA RESTRIKTIOENTSYYMIANALYYSI Molekyylibiologian ja geeniteknologian harjoitustyön kokoaminen Savoniaammattikorkeakoulun bioanalytiikan koulutusohjelmalle Opinnäytetyö Maija Vuorenpää Bioanalytiikan
POIMI KEVÄTTALVEN KAMPANJAEDUT!
 POIMI KEVÄTTALVEN KAMPANJAEDUT! Yli 400 arvoisiin NEBtilauksiin hieno NEB-TIMER! Tarjoukset voimassa 30.3.2013 asti kampanjakoodilla FIN300313FIN BioNordika Finland Oy Kutomotie 18, 00380 Helsinki tel
POIMI KEVÄTTALVEN KAMPANJAEDUT! Yli 400 arvoisiin NEBtilauksiin hieno NEB-TIMER! Tarjoukset voimassa 30.3.2013 asti kampanjakoodilla FIN300313FIN BioNordika Finland Oy Kutomotie 18, 00380 Helsinki tel
TEE-SE-ITSE-ELEKTROFOREESI
 TEE-SE-ITSE-ELEKTROFOREESI Justus Mutanen (Opinkirjon työn pohjalta) BioPop-resurssikeskus, Helsingin yliopiston LUMA-keskus Työn tavoite Työn tavoitteena on tutustua elektroforeesin periaatteeseen ja
TEE-SE-ITSE-ELEKTROFOREESI Justus Mutanen (Opinkirjon työn pohjalta) BioPop-resurssikeskus, Helsingin yliopiston LUMA-keskus Työn tavoite Työn tavoitteena on tutustua elektroforeesin periaatteeseen ja
DNA:N TOISTOJAKSOJEN MÄÄRITTÄMINEN JA YKSILÖNTUNNISTUS PCR:N AVULLA. Työohjeen testaus
 DNA:N TOISTOJAKSOJEN MÄÄRITTÄMINEN JA YKSILÖNTUNNISTUS PCR:N AVULLA Työohjeen testaus Tarja Tuovinen Kehittämistehtävä Toukokuu 2010 Solu- ja molekyylibiologian erikoistumisopinnot Tampereen ammattikorkeakoulu
DNA:N TOISTOJAKSOJEN MÄÄRITTÄMINEN JA YKSILÖNTUNNISTUS PCR:N AVULLA Työohjeen testaus Tarja Tuovinen Kehittämistehtävä Toukokuu 2010 Solu- ja molekyylibiologian erikoistumisopinnot Tampereen ammattikorkeakoulu
Molekyylibiologian perusmenetelmät
 2 3 Molekyylibiologian perusmenetelmät 740151P Biokemian menetelmät I 26.9.2017 Juha Kerätär / BMTK Päivän aiheet Mitä on molekyylibiologia? Mitä ovat keskeiset molekyylibiologian menetelmät ja mihin ne
2 3 Molekyylibiologian perusmenetelmät 740151P Biokemian menetelmät I 26.9.2017 Juha Kerätär / BMTK Päivän aiheet Mitä on molekyylibiologia? Mitä ovat keskeiset molekyylibiologian menetelmät ja mihin ne
Molekyylibiologian perusmenetelmät P Biokemian menetelmät I Juha Kerätär / BMTK
 Molekyylibiologian perusmenetelmät 740151P Biokemian menetelmät I 26.9.2017 Juha Kerätär / BMTK Päivän aiheet Mitä on molekyylibiologia? Mitä ovat keskeiset molekyylibiologian menetelmät ja mihin ne perustuvat?
Molekyylibiologian perusmenetelmät 740151P Biokemian menetelmät I 26.9.2017 Juha Kerätär / BMTK Päivän aiheet Mitä on molekyylibiologia? Mitä ovat keskeiset molekyylibiologian menetelmät ja mihin ne perustuvat?
Western blot menetelmän optimointi kantasoluista tuotetuille verkkokalvon. Jaana Huuki
 Western blot menetelmän optimointi kantasoluista tuotetuille verkkokalvon soluille Jaana Huuki Opinnäytetyö Tammikuu 2010 Laboratorioalan koulutusohjelma Pirkanmaan ammattikorkeakoulu 2 TIIVISTELMÄ Pirkanmaan
Western blot menetelmän optimointi kantasoluista tuotetuille verkkokalvon soluille Jaana Huuki Opinnäytetyö Tammikuu 2010 Laboratorioalan koulutusohjelma Pirkanmaan ammattikorkeakoulu 2 TIIVISTELMÄ Pirkanmaan
NaturaPura Ibérica Elokuu 10, 2009 Rua das Australias, No. 1 4705-322 Braga Portugali
 NaturaPura Ibérica Elokuu 10, 2009 Rua das Australias, No. 1 4705-322 Braga Portugali VIITATEN: IN VITRO IHOÄRSYTTÄVYYSTESTAUSRAPORTTI Oheisena NaturaPuran toimittaman 100% puuvillakangasmateriaalin in
NaturaPura Ibérica Elokuu 10, 2009 Rua das Australias, No. 1 4705-322 Braga Portugali VIITATEN: IN VITRO IHOÄRSYTTÄVYYSTESTAUSRAPORTTI Oheisena NaturaPuran toimittaman 100% puuvillakangasmateriaalin in
Proteiinikoon määritys (SDS-page)
") Proteiinikoon määritys (SDS-page) Johdanto SDS-pagen (sodium dodecyl sulfate) avulla määritetään proteiinien kokoja ja menetelmällä voidaan myös selvittää proteiininäytteen puhtautta. peitegeeli Menetelmässä
Proteiinikoon määritys (SDS-page) Johdanto SDS-pagen (sodium dodecyl sulfate) avulla määritetään proteiinien kokoja ja menetelmällä voidaan myös selvittää proteiininäytteen puhtautta. peitegeeli Menetelmässä
Transkriptiotekijä MAL:n ja tumalevyproteiinien välisten vuorovaikutusten tutkiminen immunosaostusmenetelmällä
 Jenni Kujala Transkriptiotekijä MAL:n ja tumalevyproteiinien välisten vuorovaikutusten tutkiminen immunosaostusmenetelmällä Metropolia Ammattikorkeakoulu Laboratorioanalyytikko (AMK) Laboratorioalan koulutusohjelma
Jenni Kujala Transkriptiotekijä MAL:n ja tumalevyproteiinien välisten vuorovaikutusten tutkiminen immunosaostusmenetelmällä Metropolia Ammattikorkeakoulu Laboratorioanalyytikko (AMK) Laboratorioalan koulutusohjelma
PIKORNAVIRUSGENOMIN MONISTUS RT-PCR- MENETELMÄLLÄ
 Opinnäytetyö (AMK) Bio- ja elintarviketekniikan koulutusohjelma Biotekniikka 2018 Tapio Metz PIKORNAVIRUSGENOMIN MONISTUS RT-PCR- MENETELMÄLLÄ OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio-
Opinnäytetyö (AMK) Bio- ja elintarviketekniikan koulutusohjelma Biotekniikka 2018 Tapio Metz PIKORNAVIRUSGENOMIN MONISTUS RT-PCR- MENETELMÄLLÄ OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio-
Tarja Huhta. FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA
 Tarja Huhta FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA Tarja Huhta Opinnäytetyö Kevät 2012 Laboratorioalan koulutusohjelma Oulun
Tarja Huhta FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA FUUSIOPROTEIINI Fc-ENDOSTATIININ TUOTTAMINEN NISÄKÄSSOLUSSA Tarja Huhta Opinnäytetyö Kevät 2012 Laboratorioalan koulutusohjelma Oulun
Original Elche antimicrobi TM desinfiointiaineen testaus Legionella lajeille
 Original Elche antimicrobi TM desinfiointiaineen testaus Legionella lajeille Tutkimusraportti 145606 10.3.2006 Sivu 1:6 Sisällysluettelo 1. YHTEYSTIEDOT... 3 2. TESTATTAVAT LEGIONELLA-LAJIT... 3 3. TESTAUSMENETELMÄT...
Original Elche antimicrobi TM desinfiointiaineen testaus Legionella lajeille Tutkimusraportti 145606 10.3.2006 Sivu 1:6 Sisällysluettelo 1. YHTEYSTIEDOT... 3 2. TESTATTAVAT LEGIONELLA-LAJIT... 3 3. TESTAUSMENETELMÄT...
FOSFORIPITOISUUS PESUAINEESSA
 FOSFORIPITOISUUS PESUAINEESSA KOHDERYHMÄ: Työ soveltuu yläkouluun kurssille elollinen luonto ja yhteiskunta. Lukiossa työ soveltuu parhaiten kurssille KE4. KESTO: Työ kestää n.1-2h MOTIVAATIO: Vaatteita
FOSFORIPITOISUUS PESUAINEESSA KOHDERYHMÄ: Työ soveltuu yläkouluun kurssille elollinen luonto ja yhteiskunta. Lukiossa työ soveltuu parhaiten kurssille KE4. KESTO: Työ kestää n.1-2h MOTIVAATIO: Vaatteita
RCA -MENETELMÄN HYÖDYNTÄMINEN GEENIKIRJASTOJEN KEHITTÄMISESSÄ
 Opinnäytetyö RCA -MENETELMÄN HYÖDYNTÄMINEN GEENIKIRJASTOJEN KEHITTÄMISESSÄ Marja Julin Bio- ja elintarviketekniikka 2011 TURUN TIIVISTELMÄ AMMATTIKORKEAKOULU Koulutusohjelma: Bio- ja elintarviketekniikka
Opinnäytetyö RCA -MENETELMÄN HYÖDYNTÄMINEN GEENIKIRJASTOJEN KEHITTÄMISESSÄ Marja Julin Bio- ja elintarviketekniikka 2011 TURUN TIIVISTELMÄ AMMATTIKORKEAKOULU Koulutusohjelma: Bio- ja elintarviketekniikka
TEKNIIKKA JA LIIKENNE. Laboratorioala OPINNÄYTETYÖ. VÄRIAINEET PIPETOINNIN APUNA qpcr-tekniikassa
 TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ VÄRIAINEET PIPETOINNIN APUNA qpcr-tekniikassa Työn tekijä: Sanna Uusivirta Työn ohjaaja: Tiina Soininen Työn ohjaaja: Jaakko Kurkela Työ hyväksytty:..
TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ VÄRIAINEET PIPETOINNIN APUNA qpcr-tekniikassa Työn tekijä: Sanna Uusivirta Työn ohjaaja: Tiina Soininen Työn ohjaaja: Jaakko Kurkela Työ hyväksytty:..
Biosensori-hiivasolujen kasvatus bioreaktorissa
 Biosensori-hiivasolujen kasvatus bioreaktorissa Organismin tiedot: - laji: Saccharomyces cerevisiae, kanta: BMA64-1A ominaisuudet ja mutaatiot: MAT α, ura 3-52, trp 1 2, leu 2-3_112 his3-11. ade2-1, can1-100
Biosensori-hiivasolujen kasvatus bioreaktorissa Organismin tiedot: - laji: Saccharomyces cerevisiae, kanta: BMA64-1A ominaisuudet ja mutaatiot: MAT α, ura 3-52, trp 1 2, leu 2-3_112 his3-11. ade2-1, can1-100
Itämeren sedimentin ja rautamangaanisaostumien. hajottaa raakaöljyä ja naftaleenia. Suomen ympäristökeskus
 Itämeren sedimentin ja rautamangaanisaostumien bakteerien kyky hajottaa raakaöljyä ja naftaleenia Mikrokosmoskokeet 23.7.-18.12.2012 Anna Reunamo, Pirjo Yli-Hemminki, Jari Nuutinen, Jouni Lehtoranta, Kirsten
Itämeren sedimentin ja rautamangaanisaostumien bakteerien kyky hajottaa raakaöljyä ja naftaleenia Mikrokosmoskokeet 23.7.-18.12.2012 Anna Reunamo, Pirjo Yli-Hemminki, Jari Nuutinen, Jouni Lehtoranta, Kirsten
PCR:n laadunvarmistus. Saija Hallanvuo Elintarvike- ja rehumikrobiologian tutkimusyksikkö/ Elintarvikemikrobiologiajaosto
 PCR:n laadunvarmistus Saija Hallanvuo Elintarvike- ja rehumikrobiologian tutkimusyksikkö/ Elintarvikemikrobiologiajaosto Ajankohtaista laboratoriorintamalla 11.10.2012 1 Laadunvarmistus Validointi Henkilökunnan
PCR:n laadunvarmistus Saija Hallanvuo Elintarvike- ja rehumikrobiologian tutkimusyksikkö/ Elintarvikemikrobiologiajaosto Ajankohtaista laboratoriorintamalla 11.10.2012 1 Laadunvarmistus Validointi Henkilökunnan
Proteiinipuhdistus. Johdanto. www.edu.fi/biogeeni
 Proteiinipuhdistus Johdanto Proteiinien jälkikäsittely on monimuotoinen, usein myös vaikeahko ja kalliskin laji. Haastetta hommassa riittääkin jos esimerkiksi E.Colin tuottamista tuhansista proteiineista
Proteiinipuhdistus Johdanto Proteiinien jälkikäsittely on monimuotoinen, usein myös vaikeahko ja kalliskin laji. Haastetta hommassa riittääkin jos esimerkiksi E.Colin tuottamista tuhansista proteiineista
"Geenin toiminnan säätely" Moniste sivu 13
 "Geenin toiminnan säätely" Moniste sivu 13 Monisteen alussa on erittäin tärkeitä ohjeita turvallisuudesta Lukekaa sivu 5 huolellisesti ja usein Vaarat vaanivat: Palavia nesteitä ja liekkejä on joskus/usein
"Geenin toiminnan säätely" Moniste sivu 13 Monisteen alussa on erittäin tärkeitä ohjeita turvallisuudesta Lukekaa sivu 5 huolellisesti ja usein Vaarat vaanivat: Palavia nesteitä ja liekkejä on joskus/usein
CORONAVIRUSTEN ESIINTYMINEN EPIDEMIOLOGISESSA TUTKIMUSAINEISTOSSA
 CORONAVIRUSTEN ESIINTYMINEN EPIDEMIOLOGISESSA TUTKIMUSAINEISTOSSA Humaanien coronavirusten esiintyvyyden tutkiminen PCR-mentelmällä Tampereen yliopiston lääketieteen laitoksen virologian yksikössä Anna
CORONAVIRUSTEN ESIINTYMINEN EPIDEMIOLOGISESSA TUTKIMUSAINEISTOSSA Humaanien coronavirusten esiintyvyyden tutkiminen PCR-mentelmällä Tampereen yliopiston lääketieteen laitoksen virologian yksikössä Anna
TEKNIIKKA JA LIIKENNE. Laboratorioala OPINNÄYTETYÖ
 TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ YERSINIA PSEUDOTUBERCULOSIS SEROTYYPPI O:9 LIPOPOLYSAKKARIDIN O-ANTIGEENIN BIOSYNTEESIÄ OHJAAVAN GEENIKLUSTERIN KLOONAAMINEN JA SEKVENSOINTI Työn tekijä:
TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ YERSINIA PSEUDOTUBERCULOSIS SEROTYYPPI O:9 LIPOPOLYSAKKARIDIN O-ANTIGEENIN BIOSYNTEESIÄ OHJAAVAN GEENIKLUSTERIN KLOONAAMINEN JA SEKVENSOINTI Työn tekijä:
IHMISEN RINOVIRUSTEN VP4/2-GEENIALUEEN MONISTAMINEN RT-qPCR - MENETELMÄLLÄ
 Opinnäytetyö (AMK) Bio- ja elintarviketekniikka Biotekniikka 2015 Aleksi Saariranta IHMISEN RINOVIRUSTEN VP4/2-GEENIALUEEN MONISTAMINEN RT-qPCR - MENETELMÄLLÄ OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU
Opinnäytetyö (AMK) Bio- ja elintarviketekniikka Biotekniikka 2015 Aleksi Saariranta IHMISEN RINOVIRUSTEN VP4/2-GEENIALUEEN MONISTAMINEN RT-qPCR - MENETELMÄLLÄ OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU
VANTAA. Brunaberget KASVIMAKROFOSSIILITUTKIMUS 2011
 VANTAA Brunaberget KASVIMAKROFOSSIILITUTKIMUS 2011 Tutkimusraportti Mia Lempiäinen-Avci Turun yliopisto Biologian laitos Kasvimuseo / Paleoetnobotaniikan laboratorio 2011 JOHDANTO Vantaan Brunabergetissä
VANTAA Brunaberget KASVIMAKROFOSSIILITUTKIMUS 2011 Tutkimusraportti Mia Lempiäinen-Avci Turun yliopisto Biologian laitos Kasvimuseo / Paleoetnobotaniikan laboratorio 2011 JOHDANTO Vantaan Brunabergetissä
Genomin ilmentyminen Liisa Kauppi, Genomibiologian tutkimusohjelma
 Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
TEKNIIKKA JA LIIKENNE. Laboratorioala
 TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ FUUSIOPROTEIININ KLOONAAMINEN, TUOTTO JA KARAKTERISOINTI Työn tekijä: Jenni Mikkonen Työn ohjaaja: Heli Haakana Työn ohjaaja: Tiina Soininen Työ hyväksytty:..
TEKNIIKKA JA LIIKENNE Laboratorioala OPINNÄYTETYÖ FUUSIOPROTEIININ KLOONAAMINEN, TUOTTO JA KARAKTERISOINTI Työn tekijä: Jenni Mikkonen Työn ohjaaja: Heli Haakana Työn ohjaaja: Tiina Soininen Työ hyväksytty:..
AMINOTRANSAMINAASI-ENTSYYMIN PUHDISTUS JA KITEYTYS
 AMINOTRANSAMINAASI-ENTSYYMIN PUHDISTUS JA KITEYTYS Saara Sipola 2011 Oulun seudun ammattikorkeakoulu AMINOTRANSAMINAASI-ENTSYYMIN PUHDISTUS JA KITEYTYS Saara Sipola Opinnäytetyö Kevät 2011 Laboratorioalan
AMINOTRANSAMINAASI-ENTSYYMIN PUHDISTUS JA KITEYTYS Saara Sipola 2011 Oulun seudun ammattikorkeakoulu AMINOTRANSAMINAASI-ENTSYYMIN PUHDISTUS JA KITEYTYS Saara Sipola Opinnäytetyö Kevät 2011 Laboratorioalan
Ma > GENERAL PRINCIPLES OF CELL SIGNALING
 Ma 5.12. -> GENERAL PRINCIPLES OF CELL SIGNALING Cell-Surface Receptors Relay Extracellular Signals via Intracellular Signaling Pathways Some Intracellular Signaling Proteins Act as Molecular Switches
Ma 5.12. -> GENERAL PRINCIPLES OF CELL SIGNALING Cell-Surface Receptors Relay Extracellular Signals via Intracellular Signaling Pathways Some Intracellular Signaling Proteins Act as Molecular Switches
1 Tehtävät. 2 Teoria. rauta(ii)ioneiksi ja rauta(ii)ionien hapettaminen kaliumpermanganaattiliuoksella.
ioneiksi ja rauta(ii)ionien hapettaminen kaliumpermanganaattiliuoksella.") 1 Tehtävät Edellisellä työkerralla oli valmistettu rauta(ii)oksalaattia epäorgaanisen synteesin avulla. Tätä sakkaa tarkasteltiin seuraavalla kerralla. Tällä työ kerralla ensin valmistettiin kaliumpermanganaatti-
1 Tehtävät Edellisellä työkerralla oli valmistettu rauta(ii)oksalaattia epäorgaanisen synteesin avulla. Tätä sakkaa tarkasteltiin seuraavalla kerralla. Tällä työ kerralla ensin valmistettiin kaliumpermanganaatti-
HIIREN TSH- RESEPTORIGEENIN PISTEMUTAATIO
 Opinnäytetyö (AMK) Bioanalytiikan koulutusohjelma Solu- ja molekyylibiologia 2013 Jenni Tuominen HIIREN TSH- RESEPTORIGEENIN PISTEMUTAATIO Geenikonstruktin rakentaminen OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN
Opinnäytetyö (AMK) Bioanalytiikan koulutusohjelma Solu- ja molekyylibiologia 2013 Jenni Tuominen HIIREN TSH- RESEPTORIGEENIN PISTEMUTAATIO Geenikonstruktin rakentaminen OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN
Flavobakteereita infektoivan ssdna-bakteriofagi V155:n karakterisointi
 Flavobakteereita infektoivan ssdna-bakteriofagi V155:n karakterisointi Jenni Marjakangas Pro gradu -tutkielma Bio- ja ympäristötieteiden laitos Solu- ja molekyylibiologia 16.9.2014 ALKUSANAT Pro gradu
Flavobakteereita infektoivan ssdna-bakteriofagi V155:n karakterisointi Jenni Marjakangas Pro gradu -tutkielma Bio- ja ympäristötieteiden laitos Solu- ja molekyylibiologia 16.9.2014 ALKUSANAT Pro gradu
Supporting Information for
 Supporting Information for Analysis of Sogatella furcifera proteome that interact with P10 protein of Southern rice black-streaked dwarf virus Win Than*, Faliang Qin*, Wenwen Liu, Xifeng Wang ** State
Supporting Information for Analysis of Sogatella furcifera proteome that interact with P10 protein of Southern rice black-streaked dwarf virus Win Than*, Faliang Qin*, Wenwen Liu, Xifeng Wang ** State
Elämän synty. Matti Leisola
 Elämän synty Matti Leisola Selitettävää Universumin rakenne Biologinen elämä Maailmallemme on olemassa kaksi erilaista selitysmallia Kaikki on syntynyt sattumanvaraisten fysikaalisten ja kemiallisten tapahtumien
Elämän synty Matti Leisola Selitettävää Universumin rakenne Biologinen elämä Maailmallemme on olemassa kaksi erilaista selitysmallia Kaikki on syntynyt sattumanvaraisten fysikaalisten ja kemiallisten tapahtumien
5 LIUOKSEN PITOISUUS Lisätehtävät
 LIUOKSEN PITOISUUS Lisätehtävät Esimerkki 1. a) 100 ml:ssa suolaista merivettä on keskimäärin 2,7 g NaCl:a. Mikä on meriveden NaCl-pitoisuus ilmoitettuna molaarisuutena? b) Suolaisen meriveden MgCl 2 -pitoisuus
LIUOKSEN PITOISUUS Lisätehtävät Esimerkki 1. a) 100 ml:ssa suolaista merivettä on keskimäärin 2,7 g NaCl:a. Mikä on meriveden NaCl-pitoisuus ilmoitettuna molaarisuutena? b) Suolaisen meriveden MgCl 2 -pitoisuus
Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa
 Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa Maria Valkonen, Kaisa Jalkanen, Martin Täubel, Anne Hyvärinen 31.3.2014 Sisäilmastoseminaari 2014 1 Tausta Asumisterveysoppaan mukaiset sisäympäristön
Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa Maria Valkonen, Kaisa Jalkanen, Martin Täubel, Anne Hyvärinen 31.3.2014 Sisäilmastoseminaari 2014 1 Tausta Asumisterveysoppaan mukaiset sisäympäristön
Agaroosigeelielektroforeesi (AGE)
") Agaroosigeelielektroforeesi (AGE) Johdanto Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNA/RNA -jaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Agaroosigeelielektroforeesin
Agaroosigeelielektroforeesi (AGE) Johdanto Agaroosigeelielektroforeesi (AGE) on yksinkertainen ja tehokas menetelmä erikokoisten DNA/RNA -jaksojen erottamiseen, tunnistamiseen ja puhdistamiseen. Agaroosigeelielektroforeesin
Virukset Materiaalitieteiden Rakennusaineina Suomalainen Tiedeakatemia
 Virukset Materiaalitieteiden Rakennusaineina Suomalainen Tiedeakatemia Mauri Kostiainen Molekyylimateriaalit-ryhmä Teknillisen fysiikan osasto Aalto-yliopisto Virukset materiaaleina Virus on isäntäsolussa
Virukset Materiaalitieteiden Rakennusaineina Suomalainen Tiedeakatemia Mauri Kostiainen Molekyylimateriaalit-ryhmä Teknillisen fysiikan osasto Aalto-yliopisto Virukset materiaaleina Virus on isäntäsolussa
Genomi-ilmentyminen Genom expression (uttryckning) Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia
 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia") Genomi-ilmentyminen Genom expression (uttryckning) DNA RNA 7.12.2017 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia Osaamistavoitteet Lärandemål Luennon jälkeen ymmärrät pääperiaatteet
Genomi-ilmentyminen Genom expression (uttryckning) DNA RNA 7.12.2017 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia Osaamistavoitteet Lärandemål Luennon jälkeen ymmärrät pääperiaatteet
Koagulaasipositiivisten stafylokokkien määrittäminen. Pesäkelaskentatekniikka.
 Vastuuhenkilöt Hakola Satu, Sivu/sivut 1 / 5 Koagulaasipositiivisten stafylokokkien määrittäminen. Pesäkelaskentatekniikka. 1 Menetelmäviitteet ja poikkeamat ISO 6888-1:1999,/ Amd 1:2003, muunnos. (Baird-Parker
Vastuuhenkilöt Hakola Satu, Sivu/sivut 1 / 5 Koagulaasipositiivisten stafylokokkien määrittäminen. Pesäkelaskentatekniikka. 1 Menetelmäviitteet ja poikkeamat ISO 6888-1:1999,/ Amd 1:2003, muunnos. (Baird-Parker
Biologian tehtävien vastaukset ja selitykset
 Biologian tehtävien vastaukset ja selitykset Ilmainen lääkiksen harjoituspääsykoe, kevät 2017 Tehtävä 2. (20 p) A. 1. EPÄTOSI. Ks. s. 4. Menetelmää käytetään geenitekniikassa geenien muokkaamisessa. 2.
Biologian tehtävien vastaukset ja selitykset Ilmainen lääkiksen harjoituspääsykoe, kevät 2017 Tehtävä 2. (20 p) A. 1. EPÄTOSI. Ks. s. 4. Menetelmää käytetään geenitekniikassa geenien muokkaamisessa. 2.
DNA-KOETTIMIEN TESTAUS YMPÄRISTÖ- SIRUA VARTEN
 DNA-KOETTIMIEN TESTAUS YMPÄRISTÖ- SIRUA VARTEN LAHDEN AMMATTIKORKEAKOULU Tekniikan ala Ympäristöteknologian koulutusohjelma Ympäristöbiotekniikka Opinnäytetyö Syksy 2010 Venla Avelin Lahden ammattikorkeakoulu
DNA-KOETTIMIEN TESTAUS YMPÄRISTÖ- SIRUA VARTEN LAHDEN AMMATTIKORKEAKOULU Tekniikan ala Ympäristöteknologian koulutusohjelma Ympäristöbiotekniikka Opinnäytetyö Syksy 2010 Venla Avelin Lahden ammattikorkeakoulu
KALAN KIR2-KANAVIEN EKSPRESSIONOPEUDEN MÄÄRITTÄMINEN EPITOOPPIMARKKERILLA (HA-TAG) NINA KEKÄLÄINEN
 NINA KEKÄLÄINEN") KALAN KIR2-KANAVIEN EKSPRESSIONOPEUDEN MÄÄRITTÄMINEN EPITOOPPIMARKKERILLA (HA-TAG) NINA KEKÄLÄINEN Pro gradu -tutkielma Itä-Suomen yliopisto Biologian laitos 2012 ITÄ-SUOMEN YLIOPISTO Biologian laitos
KALAN KIR2-KANAVIEN EKSPRESSIONOPEUDEN MÄÄRITTÄMINEN EPITOOPPIMARKKERILLA (HA-TAG) NINA KEKÄLÄINEN Pro gradu -tutkielma Itä-Suomen yliopisto Biologian laitos 2012 ITÄ-SUOMEN YLIOPISTO Biologian laitos
Lactobacillus amylovorus -kantojen pintakerrosproteiinien kloonaus ja ilmentäminen
 Oskari Torppa Lactobacillus amylovorus -kantojen pintakerrosproteiinien kloonaus ja ilmentäminen Metropolia Ammattikorkeakoulu Laboratorioanalyytikko Laboratorioalan koulutusohjelma Opinnäytetyö 18.6.2013
Oskari Torppa Lactobacillus amylovorus -kantojen pintakerrosproteiinien kloonaus ja ilmentäminen Metropolia Ammattikorkeakoulu Laboratorioanalyytikko Laboratorioalan koulutusohjelma Opinnäytetyö 18.6.2013
Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe Tehtävä 1 Pisteet / 15
 Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
MRSA-kantojen seulonta rikastusten ja selektiivisen kiinteän kasvatusalustan avulla
 Vastuuhenkilö Nykäsenoja Suvi Sivu/sivut 1 / 6 MRSA-kantojen seulonta rikastusten ja selektiivisen kiinteän kasvatusalustan avulla 1 Menetelmäviitteet ja poikkeamat Euroopan yhteisön referenssilaboratorion
Vastuuhenkilö Nykäsenoja Suvi Sivu/sivut 1 / 6 MRSA-kantojen seulonta rikastusten ja selektiivisen kiinteän kasvatusalustan avulla 1 Menetelmäviitteet ja poikkeamat Euroopan yhteisön referenssilaboratorion
Solun tutkiminen. - Geenitekniikka
 Solun tutkiminen - Geenitekniikka Tunnin sisältö 1. Bioteknologian peruskäsitteitä 2. Hieman mikroskoopeista 3. DNA:n eristäminen, puhdistaminen ja pilkkominen 4. Geenikirjasto 5. PCR 6. Elektroforeesi
Solun tutkiminen - Geenitekniikka Tunnin sisältö 1. Bioteknologian peruskäsitteitä 2. Hieman mikroskoopeista 3. DNA:n eristäminen, puhdistaminen ja pilkkominen 4. Geenikirjasto 5. PCR 6. Elektroforeesi
FNR-ISOENTSYYMIEN YLITUOTTO LITURUOHON KLOROPLASTISSA
 Opinnäytetyö (AMK) Bio- ja elintarviketekniikka Biotekniikka 2012 Ilaf Bilal FNR-ISOENTSYYMIEN YLITUOTTO LITURUOHON KLOROPLASTISSA OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio- ja elintarviketekniikka
Opinnäytetyö (AMK) Bio- ja elintarviketekniikka Biotekniikka 2012 Ilaf Bilal FNR-ISOENTSYYMIEN YLITUOTTO LITURUOHON KLOROPLASTISSA OPINNÄYTETYÖ (AMK) TIIVISTELMÄ TURUN AMMATTIKORKEAKOULU Bio- ja elintarviketekniikka
Experimental Identification and Computational Characterization of a Novel. Extracellular Metalloproteinase Produced by Clostridium sordellii
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 207 Supplementary Information Experimental Identification and Computational Characterization of
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 207 Supplementary Information Experimental Identification and Computational Characterization of
DNA RNA proteiinit transkriptio prosessointi translaatio regulaatio
 replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi translaatio
replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi translaatio
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi
 6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
Väärin, Downin oireyhtymä johtuu ylimääräisestä kromosomista n.21 (trisomia) Geeni s. 93.
 Geeni s. 93.") 1 I) Ovatko väittämät oikein (O) vai väärin (V)? Jos väite on mielestäsi väärin, perustele se lyhyesti väittämän alla oleville riveille. O/V 1.2. Downin oireyhtymä johtuu pistemutaatista fenyylialaniinin
1 I) Ovatko väittämät oikein (O) vai väärin (V)? Jos väite on mielestäsi väärin, perustele se lyhyesti väittämän alla oleville riveille. O/V 1.2. Downin oireyhtymä johtuu pistemutaatista fenyylialaniinin
Kukan kehitystä säätelevien geenien molekyylikloonaus
 Sanna Viitanen Kukan kehitystä säätelevien geenien molekyylikloonaus Opinnäytetyö Metropolia Ammattikorkeakoulu Terveys- ja hoitoala Bioanalytiikka Opinnäytetyö 15.5.2014 Tiivistelmä Tekijä(t) Otsikko
Sanna Viitanen Kukan kehitystä säätelevien geenien molekyylikloonaus Opinnäytetyö Metropolia Ammattikorkeakoulu Terveys- ja hoitoala Bioanalytiikka Opinnäytetyö 15.5.2014 Tiivistelmä Tekijä(t) Otsikko
SOLUVILJELY- TEEMANUMEROSSA MM.
 Science whereyou are Tarjoukset voimassa 30.6.2014 asti kampanjakoodilla FIN300614FIN. Kampanjahinnat ilman arvonlisäveroa. SOLUVILJELY- TEEMANUMEROSSA MM. CST:n IF-validoidut vasta-aineet ja leimatut
Science whereyou are Tarjoukset voimassa 30.6.2014 asti kampanjakoodilla FIN300614FIN. Kampanjahinnat ilman arvonlisäveroa. SOLUVILJELY- TEEMANUMEROSSA MM. CST:n IF-validoidut vasta-aineet ja leimatut
VASTAUS 1: Yhdistä oikein
 KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
Laura Hagelin & Miia Kokkola HEMOKROMATOOSI-GEENIMUTAATION KLOONAAMINEN PCR-MENETELMÄLLÄ
 Laura Hagelin & Miia Kokkola HEMOKROMATOOSI-GEENIMUTAATION KLOONAAMINEN PCR-MENETELMÄLLÄ HEMOKROMATOOSI-GEENIMUTAATION KLOONAAMINEN PCR-MENETELMÄLLÄ Laura Hagelin & Miia Kokkola Opinnäytetyö Syksy 2013
Laura Hagelin & Miia Kokkola HEMOKROMATOOSI-GEENIMUTAATION KLOONAAMINEN PCR-MENETELMÄLLÄ HEMOKROMATOOSI-GEENIMUTAATION KLOONAAMINEN PCR-MENETELMÄLLÄ Laura Hagelin & Miia Kokkola Opinnäytetyö Syksy 2013
UUDET TEKNIIKAT SISÄYMPÄRISTÖN MIKROBIEN TOTEAMISESSA
 UUDET TEKNIIKAT SISÄYMPÄRISTÖN MIKROBIEN TOTEAMISESSA LIITU-päivä 4.5.2006 FT Helena Rintala Kansanterveyslaitos, Ympäristöterveyden osasto Mihin sisäympäristön mikrobien mittauksia tarvitaan? Rakennusten
UUDET TEKNIIKAT SISÄYMPÄRISTÖN MIKROBIEN TOTEAMISESSA LIITU-päivä 4.5.2006 FT Helena Rintala Kansanterveyslaitos, Ympäristöterveyden osasto Mihin sisäympäristön mikrobien mittauksia tarvitaan? Rakennusten
1. Nimeä kuvaan nuolien osoittamat reitit/tavat, joiden kautta haitalliset aineet voivat päästä elimistöön.
 KOTITEHTÄVÄ 1 Biokemian menetelmät I syksy 2015 Opiskelijanumero: Kotitehtäviä saa miettiä ryhminä, mutta jokainen opiskelija palauttaa oman itsenäisesti käsin kirjoitetun vastauksen tehtäviin. Kotitehtävästä
KOTITEHTÄVÄ 1 Biokemian menetelmät I syksy 2015 Opiskelijanumero: Kotitehtäviä saa miettiä ryhminä, mutta jokainen opiskelija palauttaa oman itsenäisesti käsin kirjoitetun vastauksen tehtäviin. Kotitehtävästä
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. Valitse listasta kunkin yhdisteen yleiskielessä käytettävä ei-systemaattinen nimi. (pisteet yht. 5p) a) C-vitamiini b) glukoosi c) etikkahappo d) salisyylihappo e) beta-karoteeni a. b. c. d. e. ksylitoli
1. Valitse listasta kunkin yhdisteen yleiskielessä käytettävä ei-systemaattinen nimi. (pisteet yht. 5p) a) C-vitamiini b) glukoosi c) etikkahappo d) salisyylihappo e) beta-karoteeni a. b. c. d. e. ksylitoli
ALKOHOLIT SEKAISIN TAUSTAA
 ALKOHOLIT SEKAISIN TAUSTAA Kaasukromatografia on menetelmä, jolla voidaan tutkia haihtuvia, orgaanisia yhdisteitä. Näyte syötetään tavallisesti ruiskulla injektoriin, jossa se höyrystyy ja sekoittuu inerttiin
ALKOHOLIT SEKAISIN TAUSTAA Kaasukromatografia on menetelmä, jolla voidaan tutkia haihtuvia, orgaanisia yhdisteitä. Näyte syötetään tavallisesti ruiskulla injektoriin, jossa se höyrystyy ja sekoittuu inerttiin
DNA:n informaation kulku, koostumus
 DNA:n informaation kulku, koostumus KOOSTUMUS Elävien bio-organismien koostumus. Vety, hiili, happi ja typpi muodostavat yli 99% orgaanisten molekyylien rakenneosista. Biomolekyylit voidaan pääosin jakaa
DNA:n informaation kulku, koostumus KOOSTUMUS Elävien bio-organismien koostumus. Vety, hiili, happi ja typpi muodostavat yli 99% orgaanisten molekyylien rakenneosista. Biomolekyylit voidaan pääosin jakaa
Teemu Ukonaho IHMISEN FIBRONEKTIININ TYPE III TOISTOJAKSOJEN KLOONAAMINEN REKOMBINANTIN FUUSIOPROTEIININ TUOTTAMISEKSI
 Teemu Ukonaho IHMISEN FIBRONEKTIININ TYPE III TOISTOJAKSOJEN KLOONAAMINEN REKOMBINANTIN FUUSIOPROTEIININ TUOTTAMISEKSI IHMISEN FIBRONEKTIININ TYPE III TOISTOJAKSOJEN KLOONAAMINEN REKOMBINANTIN FUUSIOPROTEIININ
Teemu Ukonaho IHMISEN FIBRONEKTIININ TYPE III TOISTOJAKSOJEN KLOONAAMINEN REKOMBINANTIN FUUSIOPROTEIININ TUOTTAMISEKSI IHMISEN FIBRONEKTIININ TYPE III TOISTOJAKSOJEN KLOONAAMINEN REKOMBINANTIN FUUSIOPROTEIININ
Proteiinituoton optimointi kuoppalevyllä
 Proteiinituoton optimointi kuoppalevyllä Johdanto Ennen ison mittakaavan proteiinituottoja kasvatusolosuhteita kannattaa optimoida. Perinteisesti näitä on tehty käsityönä, mutta solukasvatusten seuranta
Proteiinituoton optimointi kuoppalevyllä Johdanto Ennen ison mittakaavan proteiinituottoja kasvatusolosuhteita kannattaa optimoida. Perinteisesti näitä on tehty käsityönä, mutta solukasvatusten seuranta
Genomin ylläpito TIINA IMMONEN MEDICUM BIOKEMIA JA KEHITYSBIOLOGIA
 Genomin ylläpito 5.12.2017 TIINA IMMONEN MEDICUM BIOKEMIA JA KEHITYSBIOLOGIA Luennon sisältö Tuman kromosomien rakenne ja pakkautuminen Pakkautumisen säätely: histonien modifikaatiot DNA:n kahdentuminen
Genomin ylläpito 5.12.2017 TIINA IMMONEN MEDICUM BIOKEMIA JA KEHITYSBIOLOGIA Luennon sisältö Tuman kromosomien rakenne ja pakkautuminen Pakkautumisen säätely: histonien modifikaatiot DNA:n kahdentuminen
VIIKKI BIOCENTER University of Helsinki
 VIIKKI BIOCENTER University of Helsinki Biologian DNA koodi ja sen selvittäminen Petri Auvinen DNA Sequencing and Genomics Laboratory Institute of Biotechnology Kuinka solut kehittyivät? Kolmenlaisia soluja
VIIKKI BIOCENTER University of Helsinki Biologian DNA koodi ja sen selvittäminen Petri Auvinen DNA Sequencing and Genomics Laboratory Institute of Biotechnology Kuinka solut kehittyivät? Kolmenlaisia soluja
Solun perusrakenne I Solun perusrakenne. BI2 I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja
 Solun perusrakenne I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja 1. Avainsanat 2. Solut tuottavat entsyymejä katalyyteiksi 3. Entsyymien rakenne ja toiminta 4. Entsyymit vaativat toimiakseen
Solun perusrakenne I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja 1. Avainsanat 2. Solut tuottavat entsyymejä katalyyteiksi 3. Entsyymien rakenne ja toiminta 4. Entsyymit vaativat toimiakseen
Shandon Cytospin Collection Fluid Shandon Instant Eosin, Alcoholic Shandon Instant Eosin, Aqueous. Thermo ELECTRON CORPORATION
 Shandon Cytospin Collection Fluid Shandon Instant Eosin, Alcoholic Shandon Instant Eosin, Aqueous Thermo ELECTRON CORPORATION Anatomical Pathology USA Clinical Diagnostics 171 Industry Drive Pittsburgh,
Shandon Cytospin Collection Fluid Shandon Instant Eosin, Alcoholic Shandon Instant Eosin, Aqueous Thermo ELECTRON CORPORATION Anatomical Pathology USA Clinical Diagnostics 171 Industry Drive Pittsburgh,
Bioanalytiikan koulutusohjelma bioanalyytikko Opinnäytetyö 16.4.2007. Riina-Mari Holopainen Paula Väre
 Automaattisen nukleiinihappoeristyslaitteen ja reaaliaikaisen PCR-menetelmän soveltuvuus herpes simplex -virusten keskushermostoinfektiodiagnostiikkaan Bioanalytiikan koulutusohjelma bioanalyytikko Opinnäytetyö
Automaattisen nukleiinihappoeristyslaitteen ja reaaliaikaisen PCR-menetelmän soveltuvuus herpes simplex -virusten keskushermostoinfektiodiagnostiikkaan Bioanalytiikan koulutusohjelma bioanalyytikko Opinnäytetyö