Hiekoittumisen ja purokunnostusten vaikutukset metsäpurojen hajottajayhteisöjen rakenteeseen ja toimintaan. Mira Niemelä
|
|
|
- Aapo Heikki Halttunen
- 8 vuotta sitten
- Katselukertoja:
Transkriptio
1 Hiekoittumisen ja purokunnostusten vaikutukset metsäpurojen hajottajayhteisöjen rakenteeseen ja toimintaan Mira Niemelä Pro Gradu -tutkielma Biologian koulutusohjelma Huhtikuu 2016

2 Sisältö 1. JOHDANTO Hajottajayhteisöjen toiminta ja merkitys puroekosysteemissä Mikrobiyhteisöt Pohjaeläimet Häiriö virtavesissä Sedimentaatio stressitekijänä virtavesissä Virtavesien kunnostukset Tutkimuksen tarkoitus AINEISTO JA MENETELMÄT Tutkimusalue Hajotustoiminta Pohjaeläinnäytteet Ympäristömuuttujat Molekyylibiologiset menetelmät DNA:n eristäminen PCR Sekvensointi Datan klusterointi Tilastolliset analyysit TULOKSET Yhteisörakenne Mikrobiyhteisöjen taksonomia Pohjaeläinten yhteisökoostumus NMDS-yhteisöordinaatiot Lajimäärä ja lajien tasaisuus Hajotus POHDINTA JOHTOPÄÄTÖKSET KIITOKSET KIRJALLISUUS Internet-lähteet... 49

3 1. JOHDANTO Puro on rajapintaekosysteemi, jossa valuma-alue, ranta-alue ja varsinainen vesiuoma ovat tiiviissä vuorovaikutuksessa keskenään (Allan & Castillo 2007). Puroympäristöt ovat monimuotoisuuskeskittymiä, jotka tarjoavat monipuolisia elinympäristöjä niin purouomassa, veden ja rannan rajapinnassa kuin rantavyöhykkeelläkin esiintyville eliöille (Raunio ym. 2008). Metsäpurojen eliöyhteisöt ovat hyvin riippuvaisia rantavyöhykkeeltä uomaan ajautuvasta orgaanisesta materiaalista ja ravinteista, sillä niiden biologinen tuotanto perustuu pitkälti rantametsästä uomaan päätyvän materiaalin, etenkin lehtikarikkeen, hajotukseen (Webster ym. 1999). Uomaan päätyvä karkea puuaines myös muokkaa uoman elinympäristöjä, ylläpitäen monimuotoista habitaattimosaiikkia erilaisiin elinympäristöihin sopeutuneille eliöille (Wallace ym. 1996, Roni ym. 2015). Virtavesiekosysteemien ekologinen tila on ihmisen toiminnan takia merkittävästi heikentynyt, minkä vuoksi niiden monimuotoisuus ja eliöyhteisöt ovat uhattuna maailmanlaajuisesti (Dudgeon ym. 2006). Tehostunut maankäyttö horjuttaa valumaalueiden luonnollisten hydrologisten ja geomorfologisten prosessien tasapainoa, mikä aiheuttaa monia erilaisia häiriöitä vesistöissä (mm. Allan 2004). Nämä häiriöt voivat yksin tai yhdessä aiheuttaa moninaisia ja jopa peruuttamattomia muutoksia virtavesien eliöyhteisöissä ja sitä kautta heikentää virtavesiekosysteemin ekologista toimintaa. Suomessa merkittävin pienten metsäpurojen tilaa heikentävä tekijä on metsäpohjan muokkauksen ja ojituksen takia lisääntynyt kiintoainekuormitus (Vuori ym. 1996, 1998, Kenttämies ym. 2006), jonka takia monet puroekosysteemit ovat muuttuneet ja niiden eliöyhteisöt uhanalaistuneet (Raunio ym. 2008). Lainsäädännön velvoittamana virtavesissä on alettu toteuttaa vesistökunnostuksia, joiden tavoitteena on usein uoman elinympäristöjen rakenteen palauttaminen luonnontilaisen kaltaiseksi (Feld ym. 2011), nykyään enenevissä määrin veden liikkeisiin ja aineiden kulkeutumiseen liittyvän prosessidynamiikan luonnonmukaistamisen kautta (Yrjänä ym. 2011). Vesistöjen fysikaalisten ominaisuuksien palauttamisen seurauksena myös eliöyhteisöjen oletetaan palautuvan häiriöstä (mm. Palmer ym. 1997). Tässä tutkielmassani tarkastelen metsätalouden aiheuttaman kiintoainekuormituksen vaikutuksia stressitekijänä metsäpuroekosysteemien ekologiaan. Tutkin purojen hajottajayhteisöjä sekä ekosysteemin energiatalouden ja siten toiminnan kannalta tärkeää lehtikarikkeen hajotusprosessia. Lisäksi

4 arvion, onko purouomien hiekoittumista vähentävillä purokunnostuksilla ollut toivottavia ekologisia vaikutuksia. 1.1 Hajottajayhteisöjen toiminta ja merkitys puroekosysteemissä Virtavesien perustuotanto on vähäistä, minkä vuoksi ekosysteemi on riippuvainen uomaan valuma-alueelta veden mukana huuhtoutuvista orgaanisesta aineesta ja ravinteista (Allan & Castillo 2007). Etenkin pienten latvapurojen ekosysteemit, joissa uoman varjostus ja ravinteiden niukkuus rajoittavat uoman sisäistä perustuotantoa, ovat hyvin riippuvaisia ekosysteemin ulkopuolelta tulevasta energiavirrasta, erityisesti rantavyöhykkeeltä uomaan ajautuvasta lehtikarikkeesta (mm. Kuehn 2015, Webster & Benfield 1986, Webster ym. 1999, Allan & Castillo 2007). Bakteereista, sienistä ja pilkkojapohjaeläimistä muodostuva hajottajaeliöstö hyödyntää lehtikariketta ravintonaan ja on tärkeä yhdysside virtavesien ravintoketjussa vapauttaessaan kuolleeseen orgaaniseen ainekseen sidoksissa olevan energian ja ravinteet kiertämään ekosysteemissä (mm. Gessner ym. 1999, Allan & Castillo 2007). Lehtikarikkeen hajoaminen tapahtuu fysikaalisten, kemiallisten ja biologisten prosessien seurauksena. Hajotusprosessi voidaan jakaa kolmeen eri vaiheeseen: 1) liukenevien aineiden vapautuminen 2) lehtien muokkaaminen ja hajotus mikrobitoiminnan kautta 3) lehtien pilkkoutuminen pohjaeläinten toiminnan ja fyysisen kulumisen seurauksena (Webster & Benfield 1986). Hajotusprosessin eri vaiheet ovat toisistaan riippuvaisia ja tapahtuvat osittain päällekkäin (Gessner ym. 1999). Mikrobit kolonisoivat uomaan pudonneet lehdet nopeasti, ja suuri osa lehtien hajotuksesta tapahtuu mikrobitoiminnan kautta, etenkin hajoamisen alkuvaiheessa (mm. Kuehn 2015). Sieniyhteisöjen merkitys etenkin hajotuksen alkuvaiheessa on suuri, ja hajotusprosessin edetessä myös bakteerit alkavat näytellä suurempaa roolia (Kuehn 2015, Webster & Benfield 1986, Webster ym. 1999, Allan & Castillo 2007). Mikrobitoiminta muokkaa lehtikarikkeen biokemiallisia ominaisuuksia, mikä saa lehdet pehmenemään ja lehtikarike on paremmin hyödynnettävissä pilkkojapohjaeläimille ravintona (Graça 2001). Lehtikarikkeella ruokailevat pilkkojapohjaeläimet hajottavat lehtikariketta pienemmiksi palasiksi ja kiihdyttävät näin olennaisesti biologista hajoamisprosessia. Pehmennyt lehtimateriaali hajoaa myös virran ja sen mukana liikkuvan materiaalin aiheuttamalla fysikaalisen kulumisen kautta (Webster & Benfield 1986, Allan & Castillo 2007). Hajotusprosessin kautta alkuperäinen lehtimateriaali muuntautuu mikrobi- ja pohjaeläinbiomassaksi, liuenneeksi ja hienojakoiseksi orgaaniseksi aineeksi, ravinteiksi ja hiilidioksidiksi sekä muiksi epäorgaanisiksi hajoamistuotteiksi 2

5 (Gessner ym 1999, Allan & Castillo 2007), ja näin kuolleisiin lehtiin varastoitunut energia vapautuu kiertämään ekosysteemissä Mikrobiyhteisöt Mikrobien, eli bakteerien ja mikrosienten, ylläpitämiä ekosysteemiprosesseja, etenkin hajotustoimintaa, on tutkittu paljon ja mikrobien tärkeys ekosysteemin toimintojen ylläpidossa on laajasti tunnistettu (mm. Kuehn 2016, Zegling 2015, Allan & Castillo 2007). Pienen koon takia mikrobilajien, yhteisöjen ja niiden ekologian tutkiminen on ollut haasteellista (mm. Bärlocker 2016), minkä takia ympäristötekijöiden ja luonnonprosessien vaikutuksista mikrobiyhteisöihin, niiden monimuotoisuuteen ja toimintaan tiedetään verrattain vähän (Heino ym. 2014). Pienen koon ja nopean elinkierron ansiosta mikrobiyhteisöt voivat olla kuitenkin herkkiä elinympäristön muutokselle ja ilmentää niitä nopeasti (Zegling 2015). Molekyylimenetelmien kehittyminen on mahdollistanut pienten organismien monimuotoisuuden ja yhteisöjen paremman tunnistamisen ja tutkimisen (Bärlocher 2010), ja mikrobiyhteisöjä ja niihin vaikuttavia ympäristötekijöitä sekä prosesseja onkin alettu tutkia ahkerasti (Bärlocher 2016, Zegling 2015). Viimeaikainen tutkimus on osoittanut, että ympäristöolosuhteet, etenkin vesikemialliset tekijät, kuten myös spatiaaliset prosessit (Heino ym. 2014), esimerkiksi lajien levittäytymisominaisuudet, sekä valuma-alueen ominaisuudet (Bergfur & Sundberg 2014) vaikuttavat mikrobiyhteisöjen paikalliseen koostumukseen. Siten ihmisperäiset muutokset veden laadussa voivat aiheuttaa muutoksia myös mikrobiyhteisöissä ja niiden toiminnassa. Mikrobiyhteisöissä onkin havaittu vasteita vesikemiallisen ympäristön, kuten lämpötilan, ph:n, happipitoisuuden sekä ravinteiden ja saasteiden määrän, muutoksiin (Bärlocher 2016, Kuehn 2016, Zegling 2015, Lecerf & Chauvet 2008) Pohjaeläimet Pohjaeläinyhteisöjä on yleisesti käytetty vesistöjen tilan tutkimiseen, sillä pitkän tutkimustradition ansiosta niiden ekologia tunnetaan verrattain hyvin ja niiden tiedetään olevan herkkiä ympäristön muutoksille (mm. Jones ym. 2012). Pohjaeläimiksi luetaan kaikki selkärangattomat eläimet, jotka elävät ainakin osan elinkierrostaan pohjasedimentissä tai sen pinnalla. Pohjaeläimiä tavataan kaikkialla virtavesissä, ja niiden monimuotoisuus on suuri (Wallace & Webster 1996). Eri lajit ovat sopeutuneet ekologialtaan erilaisiin ympäristöolosuhteisiin (Jones ym. 2012), ja paikalliset pohjaeläinyhteisöt muotoutuvat alueellisen lajipoolin ja paikallisten ympäristötekijöiden, kuten pohjan laadun ja virtausolosuhteiden, vaikutuksesta (Poff 1997, Richards ym. 1997). 3
. Pienen koon takia mikrobilajien, yhteisöjen ja niiden ekologian tutkiminen on ollut haasteellista (mm.")
6 Pohjaeläimet voidaan luokitella toiminnallisiin ryhmiin niiden ravinnonhankintatavan perusteella (Cummings 1973, Wallace & Webster 1996). Pilkkojat käyttävät ravinnokseen karkeajakoista orgaanista ainesta, kuten lehtikariketta. Laiduntajat käyttävät ravintonaan erilaisilla pinnoilla kasvavaa levää ja siihen kiinnittyneitä mikrobeja. Suodattajien ravinto koostuu pääosin hienojakoisesta orgaanisesta aineksesta, jota ne suodattavat ohi virtaavasta vedestä. Pohjakerääjät käyttävät ravintonaan pohjalle laskeutunutta hienojakoista orgaanista ainesta. Pedot käyttävät ravintonaan muita eläimiä. Virtavesien pohjaeläinlajisto muuttuu puro- ja jokiuomastoa alavirtaan edettäessä, sillä ravinnoksi käytettävän ravinnon laatu muuttuu. Latvapuroissa, missä on tarjolla runsaasti lehtikariketta, pohjaeläinyhteisössä on runsaasti pilkkojia. Hienojakoisen orgaanisen aineksen ja pohjalevien määrä kasvaa siirryttäessä jokisysteemin suistoa kohti, jolloin suodattajien, laiduntajien ja pohjakerääjien osuus lajistosta kasvaa (Vannote ym. 1980, Allan & Castillo 2007). 1.2 Häiriö virtavesissä Häiriö, eli stressi, määritellään elinympäristöön kohdistuvaksi muutokseksi, jossa jonkin ympäristötekijän vaihtelu ylittää ekosysteemin tavanomaisen vaihtelun rajat. Stressitekijän vaikutus voi aiheuttaa suoria muutoksia eliöyhteisöissä tai muutoksia esimerkiksi elinympäristön rakenteessa ja resurssien saatavuudessa, mikä taas heijastuu eliöyhteisön koostumukseen (Pickett & White 1985). Häiriön voimakkuudesta ja laadusta riippuen muutokset voivat olla hetkellisiä tai pysyviä (Lake 2000). Virtavesien eliöyhteisöt ovat sopeutuneet sietämään säännöllistä häiriötä ja palautuvat häiriön jälkeen yleensä nopeasti (mm. Resh ym 1988). Kuitenkin häiriön pidentyessä tai epäsäännöllisesti ja jatkuvasti toistuessa yhteisöt eivät välttämättä ehdi tai kykene palautumaan häiriöstä, jolloin eliöyhteisöissä ja niiden toiminnassa voi tapahtua pysyviä muutoksia (Lake 2000). Virtavedet ovat luonnostaan häiriöalttiita ekosysteemejä, ja häiriöllä on keskeinen rooli virtavesien elinympäristöjen ja eliöyhteisöjen muokkaajana (Lake 2000). Tyypillisiä luonnollisia häiriöitä ovat esimerkiksi tulvat (Poff ym. 1997), jotka syntyvät, kun valunta ympäristöstä kasvaa merkittävästi esimerkiksi lumen sulamisen tai kasvaneiden sademäärien takia (Allan & Castillo 2007). Suuret vesimassat uurtavat ja kuluttavat jokiuomaa, tempaavat mukaansa uoman pohjalle kasaantunutta hienojakoista ainesta, vesikasveja ja muuta materiaalia sekä siirtävät suurempiakin kiviä, joten tulvat muokkaavat suuresti uoman pohjan rakennetta, tuhoten ja samalla luoden elinympäristölaikkuja (Poff ym. 1997, Lake 2000). Veden mukana uomiin kulkeutuu myös maa-ainesta, joka kulkeutuu virran mukana tai kasautuu ja tiivistyy uoman pohjalle. Äärivirtaamat ja sedimentaatio ovat pääasialliset 4

7 häiriötä aiheuttavat stressitekijät virtavesissä (Poff ym. 1997, Wood & Armitage 1997). Veden liikkeisiin ja sedimentin kulkeutumiseen liittyvät hydrologiset ja geomorfologiset prosessit ovat tärkeitä jokiuoman rakennetta muovaavia tekijöitä, jotka luovat sekä paikallisesti että ajallisesti vaihtelevat ympäristöolosuhteet ja tekevät virtavesistä luonnostaan dynaamisia ekosysteemejä (mm. Elosegi ym. 2010) Sedimentaatio stressitekijänä virtavesissä Kiintoainetta päätyy vesistöihin luonnostaan niin sanotun luonnonhuuhtouman kautta, millä on tärkeä rooli purojen elinympäristö- ja ravintoresurssien ylläpidossa (Wood & Armitage 1997, Kemp ym. 2011, Elosegi ym. 2010). Vapaa vesi kulkeutuu jokeen sitä ympäröivältä valuma-alueelta pinta- ja pintakerrosvaluntana maaperässä sekä pohjavesivaluntana kosketuksissa kallioperään, ja veden mukana kulkeutuu ravinteita, sedimenttejä sekä orgaanista ainesta jokiuomaan (mm. Poff ym. 1997, Allan & Castillo 2007). Veden ja kiintoaineiden kulkeutumista valuma-alueella säätelevät alueen ilmasto, kallio- ja maaperän pinnanmuodot, maaperän ominaisuudet sekä kasvillisuus (Allan & Castillo 2007). Luonnontilaisilla valuma-alueilla kiintoaineen kulkeutuminen luonnonhuuhtoumana on yleensä vähäistä, sillä häiriintymättömässä metsäekosysteemissä kasvillisuus vakauttaa maaperää sitoen maa-ainesta ja vettä, näin ehkäisten eroosiota (Ahtilainen & Huttunen 1995). Viimeisten vuosikymmenten aikana lisääntynyt ja tehostunut maankäyttö, kuten maatalous, metsätalous ja rakentaminen, on muovannut merkittävästi valuma-alueiden metsiä ja suoalueita (mm. Allan 2004, Allan & Castillo 2007). Esimerkiksi Suomessa suuri osa metsäalasta on metsätalouden piirissä, ja valtaosa turvemaista on ojitettu (Ahvonen ym. 1992, Kenttämies ym. 2006). Laaja-alaisilla ojituksilla on pyritty kuivattamaan metsänpohjaa ja siten tehostamaan metsänkasvua (Peltomaa 2007). Metsätaloudellisesti muokattujen valuma-alueiden valunta nopeutuu ja kasvaa, sillä maan vedenpidätyskyky heikkenee muuan muassa hakkuiden muokatessa kasvillisuutta, ja suuri osa sadevedestä päätyy metsäojiin ja niistä edelleen virtavesiin (mm. Ahvonen ym. 1992, Kenttämies ym. 2006, Allan & Castillo 2007), ja näin myös vesistöihin kulkeutuvan kiintoaineksen määrä kasvaa. Lisääntynyt valunta kasvattaa myöskin virtaamaa ja sen vaihtelua, mikä vaikuttaa uoman eroosioon ja kiintoaineksen käyttäytymiseen uomassa (Wood & Armitage 1997). Kiintoainekuormitukselle on ominaista voimakas ajallinen vaihtelu sääolojen ja maaperän kosteusolosuhteiden vaihdellessa. Esimerkiksi kuivilla sääjaksoilla huuhtouma on vähäistä, 5
. 1.2.")
8 kun taas tulva-aikoina ja rankkojen sateiden aikaan aineiden kulkeutuminen lisääntyy (mm. Wood & Armitage 1997, Kenttämies ym. 2006). Kasvanut kiintoainekuormitus muokkaa jokiuoman fysikaalista rakennetta, ja muuttaa näin uomassa elävien eliöiden elinympäristöjä (Wood & Armitage 1997, Owens ym. 2005). Pohjan keskimääräinen raekoko pienenee ja kivet sekä soraikot peittyvät tai niiden välitilat täyttyvät hiekalla (Wood & Armitage 1997). Suorilla ja hitaasti virtaavilla puro-osuuksilla uoma voi kokonaan täyttyä hiekasta, mikä vähentää uoman luonnollista syvyysvaihtelua. Hiekkakasaumat vaikuttavat uoman paikallisiin virtausolosuhteisiin, ja virtausnopeudeltaan erilaiset habitaatit uomassa vähenevät (Wood & Armitage 1997, Kemp ym. 2011). Hienojakoinen hiekka myös liikkuu helpommin virran mukana kuin karkeampi aines (Newbury 1984), joten pohjan vakaus vähenee sekä muuan muassa virtaamahäiriön ulottumattomissa olevat olosuhteiltaan vakaat pohjaeläinten turvapaikat vähenevät (Jones ym. 2012). Nämä muutokset vaikuttavat uomassa eliöille tarjolla olevien habitaattien laatuun ja heterogeenisyyteen sekä lisäävät uoman pohjan häiriöherkkyyttä, ja voivat aiheuttaa myös muutoksia eliöyhteisöissä. Erityisesti pohjaeläimet ovat sopeutuneet elämään virrannopeudeltaan ja pohjan koostumukseltaan tietynlaisissa habitaateissa, joten pohjan hiekoittumisella on vaikutusta etenkin pohjaeläinyhteisöjen rakenteeseen (Jones ym. 2012, Richards ym. 1997) (kuva 1). Koska uoman fysikaalisen rakenteen monimuotoisuus ylläpitää biologista monimuotoisuutta (Muotka & Syrjänen 2007), voi hiekoittumisen myötä yksipuolistunut uoman rakenne myös alentaa yhteisön lajirikkautta. Hiekkakuormitus voi vaikuttaa paitsi elinympäristön rakenteeseen ja laatuun, myös saatavilla olevien ravintoresurssien määrään ja laatuun (kuva 1). Koska hiekka kulkeutuu helposti veden mukana, hiekoittuminen lisää uoman epävakautta. Pilkkojapohjaeläinten ja mikrobien ravintona käyttämä lehtikarike voi hautautua hiekkaan tai kulkeutua virran mukana muualle (Muotka & Laasonen 2002, Lepori ym. 2005a), jolloin lehtikarike ei ole resurssina käytettävissä tai sen käytettävyys on heikentynyt. Pohjan epävakaus ja veden lisääntynyt sameus myös vaikeuttaa leväbiofilmien muodostumista uoman pohjalle, ja aiheuttaa näin muutoksia leviä ravintona hyödyntävien laiduntajien ravintoresurssiin (Kemp ym. 2011, Jones ym. 2012). Lisääntyneen valunnan kautta hienojakoisen orgaanisen aineksen kulkeutuminen uomaan voi lisääntyä, jolloin suodattajapohjaeläimet voivat hyötyä (Jones ym. 2012). 6

9 Hiekkakuorma voi vaikuttaa lehtikarikkeen hajoamiseen, sillä lehtien hautautuessa hiekkaan pilkkojapohjaeläinten sekä mikrobien pääsy karikkeeseen vaikeutuu, mikä voi hidastaa hajoamista (Benfield ym. 2001, Danger ym. 2012). Paksu sedimenttikerros myös vähentää lehtien fyysistä kulumista, kun virtauksen kuluttavat voimat eivät pääse vaikuttamaan lehtikarikkeeseen (Cornut ym. 2010, Danger ym. 2012). Toisaalta hiekoittuminen voi nopeuttaa lehtikarikkeen hajoamista, sillä veden mukana liikkuva kiintoaines voi lisätä lehtikarikkeen fyysistä kulumista, mikäli karike ei peity eristävällä hiekkakerroksella (Matthaei ym. 2010). Kuva 1. Havainnekuva lisääntyneen kiintoainekuormituksen negatiivisista vaikutuksista puroekosysteemin fysikaalisessa rakenteessa ja mekanismit pohjaeläinten yhteisömuutoksen takana (laadittu Kemp ym pohjalta). *BOD = biologien hapenkulutus. Nelikulmiot kuvaavat fysikaalisia muutoksia ja ovaalit biologisisia muutoksia sekä vasteita ympäristömuutokselle. 7

10 1.3 Virtavesien kunnostukset Virtavesien ekologisen tilan heikentymiseen on herätty myös yhteiskunnallisesti. Euroopan unionin vesipuitedirektiivi (2000/60/EY) ja siihen perustuva Suomen vesienhoitolaki (1299/2004) edellyttävät pintavesien ekologisen tilan arviointia ja seurantaa. Seurannan painopistettä on haluttu kehittää kohti kokonaisvaltaisempaa vesien tilan seurantaa ja suojelua, ja arvioinnissa pyritään tunnistamaan ennen kaikkea ihmistoiminnan vaikutuksia eliöyhteisöihin ja ekosysteemin toimintakykyyn (Hering ym. 2011). Ensisijaisia tavoitteita ovat vesiekosysteemien toiminnan säilyttäminen, biologisen monimuotoisuuden turvaaminen sekä kestävä, monipuolisen vesivarantojen käytön mahdollistava vesien hoito (Euroopan parlamentti ja neuvosto 2000, Apitz ym. 2006). Päämääränä on ollut saavuttaa vesistöjen hyvä ekologinen tila pääosin vuoteen 2015 mennessä, ja lopullisesti tavoite tulisi saavuttaa vuoteen 2027 mennessä. Lainsäädäntö velvoittaa vesistön tilan parantamiseen siellä, missä se on ihmistoiminnan seurauksena merkittävästi huonontunut, mikä on johtanut laajamittaiseen vesistöjen kunnostamiseen ja kunnostusten ekologisten vaikutusten tutkimiseen (Feld ym. 2011). Vesistökunnostusten kautta vesistöjen fysikaalisia ominaisuuksia pyritään palauttamaan luonnontilaisen kaltaisiksi (Feld ym. 2011), minkä kautta myös eliöyhteisöjen odotetaan elpyvän häiriöstä (Palmer ym. 1997). Eri mittakaavan jokikunnostuksia on toteutettu luvulta lähtien, ja kunnostukset ovat perustuneet usein sille oletukselle, että habitaattien heterogeenisyys edistää biologista monimuotoisuutta (Lepori ym. 2005b, Palmer ym. 1997, Feld ym. 2011). Käytännön kunnostustoimet ovat keskittyneet uoman rakenteelliseen elinympäristöjen kunnostamiseen, joissa on pyritty uoman habitaattien ja virtausolojen monipuolistamiseen ja sitä kautta biologisen monimuotoisuuden lisäämiseen (Feld ym. 2011). Näillä toimilla on ollut usein lyhytkantoisia, heikkoja ja ristiriitaisia vaikutuksia biologisiin yhteisöihin (mm. Palmer ym. 2010), sillä kunnostustoimet ovat olleet monissa tapauksissa mittakaavaltaan riittämättömiä hallitsemaan suuremman mittakaavan häiriöprosessia (Feld ym. 2011). Esimerkiksi hiekoittuneen uoman habitaattien monimuotoisuutta lisäävillä uoman sisäisillä kunnostustoimilla on vain lyhytaikainen vaikutus, jos valuma-alueen suuremman mittakaavan hydrologisten ja geomorfologisten prosessien häiriötä ei saada hallittua ja hienojakoisen aineksen kulkeutuminen valumaalueelta uomaan jatkuu (mm. Roni ym. 2008, Lorenz & Feld 2013). Viime vuosina kunnostusekologiassa on siirrytty elinympäristöjen kunnostamiseen prosessiajattelun kautta, 8
.")
11 missä häiriintynyt prosessi, esimerkiksi sedimentaatio ja sen myötä vahingoittunut elinympäristö, pyritään palauttamaan eri mittakaavan kunnostustoimilla luonnontilaan (mm. Feld ym. 2011). Hydrologiset, geomorfologiset ja biologiset prosessit vaikuttavat toinen toisiinsa, ja näiden vuorovaikutusten mekanismeja ei vielä täysin ymmärretä (Vaughan ym. 2009). Biologisten yhteisöjen heikot vasteet kunnostustoimiin johtuvat luultavasti siitä, ettemme vielä täysin ymmärrä todellisia biologisia yhteisöjä ohjaavia sekä määrittäviä tekijöitä ja prosesseja (Palmer ym. 2010, Roni ym. 2008). Esimerkiksi biologisten vuorovaikutusten merkitys eliöyhteisöjen muokkaajana on vielä heikosti ymmärretty (Feld ym. 2011), mikä tekee yhteisötason vaikutusten ennustamisesta erityisen haasteellista. 1.4 Tutkimuksen tarkoitus Tutkimuksen tarkoituksena on selvittää metsätaloustoimien, lähinnä metsäojitusten, aiheuttaman sedimentaatio- eli hiekkakuormituksen merkitystä stressitekijänä pienissä metsäpuroissa. Tarkastelukohteena ovat purojen hajottajayhteisöt, eli sienet, bakteerit ja pohjaeläimet, sekä niiden ylläpitämä hajotusprosessi, joiden kautta arvioidaan, onko purouomien hiekoittuminen vaikuttanut puroekosysteemien yhteisörakenteeseen ja ekologiseen toimintakykyyn. Tutkimuksessa analysoidaan eri muuttujien roolia yhteisöjä määrittävinä ja hajotusprosessiin vaikuttavina tekijöinä. Lisäksi tutkimuksessa tarkastellaan, onko sedimentaatiokuormituksen vähentämiseen pyrkivillä purokunnostuksilla ollut toivottavia ekologisia vaikutuksia. Tutkimuksessa vertaillaan sedimentaatiosta kärsiviä kunnostettuja ja kunnostamattomia puroja lähes luonnontilaisiin vertailupuroihin. Tutkimuksessa oletetaan, että hiekoittumisen seurauksena nähdään vasteita pohjaeläinyhteisöjen rakenteessa, sillä sedimentaatiokuormitus vaikuttaa pohjaeläinten elinympäristön rakenteeseen sekä saatavilla olevien ravintoresurssien laatuun. Koska hiekoittuminen ei juuri vaikuta veden laatuun, sieni- ja bakteeriyhteisöissä ei oleteta nähtävän suuria vasteita. Lisäksi hajotusprosessin oletetaan hidastuvan hiekoittumisen seurauksena. 9
. Esimerkiksi biologisten vuorovaikutusten merkitys eliöyhteisöjen muokkaajana on vielä heikosti ymmärretty (Feld ym.")
12 2. AINEISTO JA MENETELMÄT 2.1 Tutkimusalue Tutkielmani on osa Suomen ympäristökeskuksen ja Oulun yliopiston tutkimusprojektia (ReFFECT, Restoration Effectiveness of Sediment Stressed Forest Streams), jossa selvitetään metsätaloudesta aiheutuvan sedimenttikuormituksen vaikutusta stressitekijänä pienten metsäpurojen ekologiaan, sekä sedimenttikuormitusta vähentävien jokikunnostusten ekologista onnistumista. Tutkimuspurot sijaitsevat Koillismaalla Iijoen valuma-alueella (Kuva 2), mistä aineisto on kerätty pääosin syksyllä Purot on pyritty valitsemaan niin, että ne olisivat kooltaan mahdollisimman samankaltaisia. Tutkimukseen valittiin yhteensä 32 kohdetta: 11 vertailupuroa, 12 sedimentaatiosta kärsivää puroa ja 9 kunnostettua puroa. Kunnostetut purot valittiin Metsähallituksen vuosina kunnostamista puroista, jotka kärsivät metsäojitusten ja metsäautoteiden aiheuttamasta hiekoittumisesta. Kunnostukset on toteutettu sekä uoma- että valuma-aluekunnostustoimenpiteillä (Yrjänä ym. 2011). Valuma-aluetoimien (metsäojien tukkiminen, pintavaluntakenttien ja laskeutusaltaiden rakentaminen, soiden ennallistaminen) tarkoituksena on vähentää kiintoaineen kulkeutumista valuma-alueelta uomaan ja palauttaa purojen luontainen hydrologia. Uoman sisäisillä kunnostustoimilla (kiveäminen, sorastaminen, puiset altakaivaja-, virranohjaus- ja tulvitusrakenteet, vanhan uoman vesitys) pyritään ennallistamaan ja monipuolistamaan uoman fysikaalista rakennetta ja luomaan olosuhteet, joiden kautta uoman pohja lajittuu ja puhdistuu liiallisesta hiekasta luonnollisten prosessien kautta (Yrjänä ym. 2011). Kunnostusten ekologista onnistumista tarkasteltiin vertailemalla kunnostettuja puroja joukkoon hiekoittuneita ja lähes luonnontilaisia puroja (paine-kontrollivertailuasetelma). Vertailupurojen valinta perustui valuma-alueiden maankäytön voimakkuutta kuvaavaan turvemaiden ojitusprosenttiin. Lähes luonnontilaisten vertailupurojen joukkoon pyrittiin valitsemaan puroja, joiden valuma-alue oli ojittamaton tai mahdollisimman vähän ojitettu. Vastaavasti hiekoittumisesta kärsivien purojen joukkoon valittiin valuma-alueeltaan vahvasti ojitettuja puroja, joissa oli selviä merkkejä luontaisen hiekan määrän ylittävästä hiekoittumisen tasosta. Tarkemmat purokohtaiset tiedot löytyvät liitteestä 1. 10
, mistä aineisto on kerätty pääosin syksyllä 2013.")
13 Kuva 2. Tutkimuspurojen sijainti Iijoen valuma-alueella. Tutkimuspaikat on merkitty pisteillä. (IMP=hiekoittunut puro, REF=vertailupuro, REST=kunnostettu puro).

14 2.2 Hajotustoiminta Hajotustoimintaa havainnoitiin tutkimusuomiin sijoitettujen lehtikarikepussien avulla (Boulton & Boon 1991). Pussien sisään suljettiin kuusi grammaa harmaalepän (Alnus incana) lehtiä, jotka kerättiin Oulun Taskisenperän venesataman alueelta (N E ) elokuussa Lehtien annettiin kuivua huoneenlämmössä noin kaksi viikkoa ennen pussittamista. Tutkimuksessa käytettiin sekä harvoja (5 kpl/uoma, silmäkoko 8 mm) että tiheitä (5 kpl/uoma, silmäkoko 0,2 mm) pusseja. Pohjaeläimet eivät pääse hajottamaan tiheiden pussien lehtikariketta, joten vertailemalla harvojen pussien hajotusta tiheisiin pusseihin pystyttiin arvioimaan pohjaeläinten ja mikrobien osuutta lehtien kokonaishajotuksesta (Hieber & Gessner 2002). Jokaiseen tutkimusuomaan sijoitettiin kymmenen lehtikarikepussia siten, että viiteen satunnaisesti valittuun paikkaan uomassa sijoitettiin sekä harva että tiheä pussi vierekkäin. Pussit ankkuroitiin uoman pohjaan rakennustiilien avulla. Lehtipussit sijoitettiin uomiin syyskuun puolessa välissä lehtien putoamisen aikaan. Noin kolmen viikon kuluttua pussit kerättiin uomista, suljettiin muovipusseihin ja pakastettiin jatkokäsittelyä varten. Laboratoriossa hieman sulaneesta lehtimateriaalista otettiin omenaporalla DNA- sekä ergosterolinäytteet, jotka sijoitettiin pieniin paperipusseihin, kuivattiin yön yli pakastekylmäkuivurilla -55 celsiusasteessa ja punnittiin. Harvoista lehtikarikepusseista kerättiin pohjaeläimet, jotka säilöttiin 70 % etanoliin lajinmääritystä varten. Jäljelle jäänyt alkuperäinen lehtimateriaali eroteltiin muusta materiaalista paperipusseihin. Kuivapainon määrittämistä varten lehtimateriaalin annettiin kuivua lämpöuunissa 60 asteessa noin vuorokauden ajan. Punnitut lehtinäytteet tuhkattiin polttouunissa 550 asteessa neljä tuntia, jolloin saatiin määritetty tuhkan eli epäorgaanisen materiaalin paino. Lehtikarikkeen hajoamisen oletettiin noudattavan eksponentiaalista hajoamismallia. Täten jokaiselle lehtipussille määritettiin lehtikarikkeen hajoamisnopeus (k) yhtälöstä Mr = Mi * e -kd, missä Mr on uiton jälkeen jäljelle jäänyt tuhkaton lehtimassa, Mi on lehtien kuivamassa ennen uittoa ja d uittopäivien lukumäärä (Benfield 1996). Jokaisessa uomassa yhteen lehtipussiin laitettiin lämpötilankirjausanturi tarkkailemaan tutkimusajan lämpötilaa. Hajoamisnopeus korjattiin puron veden vuorokauden keskilämpötilojen lämpösummalla, sillä hajoamisnopeuden on todettu olevan lämpötilasta riippuvainen (Webster & Benfield 1986).

15 2.3. Pohjaeläinnäytteet Puroista (n=29) kerättiin pohjaeläinaineisto standardoidulla menetelmällä (Meissner ym. 2013). Jokaisesta uomasta otettiin noin metrin matkalta neljältä paikalta 30 sekunnin potkuhaavinäyte siten, että jokaisen näytealan kattavuus oli noin yksi neliömetri näytettä kohden. Näytepaikat pyrittiin valitsemaan satunnaisesti, kuitenkin niin että paikat edustavat koskimaista habitaattia. Näytteet säilöttiin 70-prosenttiseen etanoliliuokseen ja siirrettiin laboratorioon poimintaa ja määritystä varten. Laboratoriossa pohjaeläimet eroteltiin näytteistä muun aineksen seasta ja määritettiin ympäristöhallinnon tavoitetaksonomian määritystasolle (Meissner ym. 2013), pääosin lajitasolle. Pohjaeläimet jaoteltiin neljään eri ravinnonkäyttöryhmään perustuen Heinon & Juntusen (2001) sekä Moog:n (2002) jaotteluun: pilkkojat, laiduntajat, suodattajat ja pedot. Tarkastelusta jätettiin pois pohjakerääjät, joiden osuus aineistosta oli hyvin pieni, sekä ryhmät, jotka sisältävät useita eri ravinnoinkäyttöryhmiä, kuten surviaissääsket Chironomidae Ympäristömuuttujat Samaan aikaan pohjaeläinnäytteenoton kanssa tutkimuspaikoilta kartoitettiin uoman habitaattia, vesikemiaa ja fysikaalisia ominaisuuksia kuvaavia ympäristömuuttujia. Tutkimuspaikoilta kirjattiin itä- ja pohjoiskoordinaatit. Jokaisen lehtipussin päältä mitattiin sekä tutkimuksen alku- että lopputilanteessa ( ) virrannopeus sekä veden syvyys. Uoman leveys ja kaltevuus arvioitiin viideltä kohtaa uomasta, sekä uoman varjostus 20 kohtaa uoman keskeltä (latvuksen prosenttipeittävyys). Arviointien perusteella laskettiin uomakohtaiset keskiarvot. Jokaisesta purosta otettiin vesinäyte, josta määritettiin laboratoriossa veden väriluku, kokonaisfosforipitoisuus sekä kiintoaineksen määrä. Sähkönjohtavuus ja ph mitattiin maastossa YSI Professional plus-mittarilla ( YSI). Uoman pohjan prosentuaalinen sammalpeittävyys ja raekoko arvioitiin silmämääräisesti 15 tutkimusjaksolle satunnaisesti sijoitetulta 50 x 50 cm ruudulta. Raekoon määrittämisessä käytettiin Wentworthin (1922) yhdeksänportaista asteikkoa, joka vaihtelee siltistä ja hiekasta (halkaisija 0,25 2 mm) suuriin lohkareisiin ja kallioperään (halkaisija >400 mm), arvioimalla kunkin luokan prosenttipeittävyys. Arviointien perusteella laskettiin raekoon keskiarvo kullekin tutkimusjaksolle. Purojen karkean puuaineksen määrä (m 3 /m 2 ) arvioitiin määrittämällä puun kuutiomäärä uomassa ja suhteuttamalla se uoman pinta-alaan tutkimusjaksolla. Jokaiselle tutkimuspaikalle määritettiin myös uoman ja sen valuma-alueen 13

16 häiriöherkkyyttä kuvaava Pfankuch-luku. Pfankuch-menetelmässä arvioidaan muuan muassa uoman pohjan ja törmän vakautta, perustuen muuan muassa pohjan laadun sekä törmän materiaalin ja kasvillisuuden arviointiin. Luku kuvastaa uoman herkkyyttä virtaamavaihtelun aiheuttamalle eroosiolle ja materiaalin kulkeutumiselle (Pfankuch 1975). Tutkimuspurojen valuma-alueen koko, korkeus merenpinnasta ja maankäyttömuuttujat selvitettiin ArcMap-ohjelmistolla Corine-maankäyttöluokitusaineistosta. 2.5 Molekyylibiologiset menetelmät DNA:n eristäminen DNA-näytteet otettiin osittain sulaneesta lehtikarikkeesta erikseen bakteeri- ja sieniyhteisöjen määrittämistä varten. Kuivaamisen ja punnitsemisen jälkeen (kts. osa 2.2) jokaisen puron viisi näytettä yhdistettiin, harvat ja tiheät pussit omaksi kokoomanäytteekseen. Kuivattu lehtimateriaali jauhettiin huhmareella. Noin 25 milligrammaa hienonnettua lehtimateriaalia käytettiin DNA:n eristetykseen, joka toteutettiin Powersoil -eristyssarjalla (MO BIO Laboratories 2013). Eristämisen onnistumista tarkasteltiin spektrofotometrisesti NanoDrop-laitteella (Thermo Scientific), jolla saadaan karkea arvio DNA:n määrästä ja puhtaudesta (Desjardins & Conklin 2010) PCR Eristetyt näytteet monistettiin polymeraasiketjureaktiossa (PCR), jossa eristetyt DNAjuosteet monistetaan moninkertaiseksi (Madigan ym. 2009). DNA:n monistus toteutettiin erityisellä PCR-laitteella (Veriti Thermal Cycler, Life Technologies). Jokaisesta DNAnäytteestä tehtiin kolme rinnakkaista näytettä PCR-reaktiota varten, jotka yhdistettiin reaktion jälkeen. Näin pyrittiin varmistamaan monistumisen onnistuminen ja minimoimaan pipetointitarkkuudesta aiheutuvat häiriöt. PCR:n onnistuminen tarkistettiin MultiNA ajolla (Microchip Electrophoresis System for DNA/RNA, Shimadzu Corporation). PCRreaktioliuoksen komponentit on esitetty taulukossa 1. Alukkeiden (A & B) tehtävänä on rajata monistettava alue DNA-juosteen 5 ja 3 - päästä, joiden mukaan DNApolymeraasientsyymi (Hot Phusion Start II) monistaa määrätyn alueen DNA-juosteesta. Bakteerinäytteille käytettiin ribosomaalisen DNA:n (rdna) 16S-alueen alukkeita 519F 5 - CAGCMGCCGCGGTAATWC-3 ja 926trP1 5 -CCTCTCTATGGGCAGTCGGTGAT- CCGTCAATTCCTTTRAGTTT-3 sekä sieninäytteille rdna:n ITS-alueen alukkeita ITS1F 5 -CTTGGTCATTTAGAGGAAGTAA-3 ja 58A2R-P1 5 - CCTCTCTATGGGCAGTCGG-TGATCTGCGTTCTTCATCGAT-3. Jokaista näytettä 14

17 varten oli oma yksilöllinen A-aluke, jonka perusteella näyte pystyttiin myöhemmin tunnistamaan. Polymeraasientsyymi rakentaa uuden juosteen dntp-molekyyleistä eli deoksinukleotideistä. Puskuriliuoksen tehtävänä (GC) on muuttaa reaktion olosuhteet otollisiksi DNA:n monistumiselle ja stabiloida polymeraasientsyymin toimintaa (Madigan ym. 2009). PCR-reaktiota varten valmistettiin ns. mastermix-liuos, joka mitoitettiin riittämään kaikille analysoitaville näytteille. Mastermix-liuoksen valmistamisessa muut komponentit lisättiin steriiliin veteen, entsyymi viimeisenä. Liuosta pipetoitiin 21 mikrolitraa yksittäistä näytettä kohden 96-kuoppalevylle. Liuokseen lisättiin kolme mikrolitraa eristettyä DNA:ta ja lopuksi yksi mikrolitra A-aluketta. Näin jokaisen PCR-näytteen reaktioliuoksen tilavuus oli 25 mikrolitraa (Taulukko 1). Taulukko 1. PCR-reaktioliuoksen valmistamiseen käytetyt komponentit. 5,0 µl GC-puskuri 0,5 µl dntp 0,5 µl B-aluke 0,5 µl Phusion Hot Start II-entsyymi 13,5 µl Steriili vesi = 21 µl Mastermix + 3 µl Eristetty DNA + 1 µl A-aluke = 25 µl Yhteensä PCR-reaktiossa toistetaan kolmea vaihetta: denaturaatio, alukkeiden liittyminen ja pidentyminen (Madigan ym. 2009). Taulukossa 2 on esitetty käyttämämme PCR-ohjelma ja sen vaiheet (katso myös Heino ym. 2015). Denaturaatiovaiheessa reaktioseos kuumennettiin 98 celsiusasteeseen 10 sekunnin ajaksi, jolloin DNA denaturoituu ja sen vastinjuosteet irtoavat toisistaan. Liittymisvaiheessa lämpötila laskettiin 64 celsiusasteeseen, jolloin alukkeet kiinnittyvät denaturoituneeseen DNA-juosteeseen emäsparisäännön mukaisesti. Alukkeiden kiinnittymisen jälkeen DNA-polymeraasi voi aloittaa toimintansa. Pidentymisvaiheessa DNA-polymeraasi luo vastinjuosteen templaatti-dna:lle. Pidentymisvaihe toteutettiin entsyymille optimaalisessa 72 celsiusasteen lämpötilassa. Tämä 15

18 vaiheiden sykli toistettiin 25 kertaa. Ennen varsinaista monistusvaihetta näytteille suoritettiin kolmen minuutin alkudenaturaatio kaikkien DNA-molekyylien denaturoitumisen varmistamiseksi. Lopuksi suoritettiin loppupidentyminen (5 minuuttia), jossa varmistettiin kaikkien juosteiden kopioituminen loppuun asti. Ohjelman lopuksi lämpötila laskettiin neljään celsiusasteeseen. Koska osa näytteistä oli vaikeasti monistuvia, toistettiin sykli näille näytteille 30 kertaa ja eristettyä DNA:ta lisättiin 5 mikrolitraa, jolloin steriilin veden määrä laskettiin 11,5 mikrolitraan näytettä kohden. Taulukko 2. PCR-ohjelman eri vaiheiden lämpötilat ja inkubaatioajat bakteeri- ja sieninäytteille (Suluissa merkitty sienten ohjelmatieto, mikäli se on eronnut bakteerien ohjelmasta). Reaktion päävaiheet Osavaihe Lämpötila ( C) Aika (m.ss) Syklien määrä Alkudenaturaatio :00 1 Denaturaatio :10 Alukkeiden kiinnittyminen 2 64 (58) 0:30 Pidentyminen :20 (0:15) Lopullinen pidentyminen : Sekvensointi Eristetty ja monistettu DNA sekvensoitiin Ion Torrent-sekvensointilaitteella (Ion Torrent TM Personal Genome Machine (PGM) System, Life Technologies), jossa hyödynnetään puolijohdesekvensointitekniikkaa. Laite muuntaa biokemiallisen informaation digitaaliseen muotoon (FASTQ) reaaliaikaisesti. Tässä menetelmässä sekvensointi perustuu DNA:n polymerisaation aikana vapautuvien vetyionien havaitsemiseen (Life Technologies Corporation 2013) Ennen sekvensointia yhdistetty DNA-näyte monistetaan emulsio-pcr:n avulla. DNA pilkotaan pieniksi fragmenteiksi, joiden päihin liitetään adapterit. Adaptereiden avulla fragmentit kiinnittyvät helmiin, joiden pinnalla DNA:n monistuminen tapahtuu. Yhteen helmeen voi kiinnittyä vain yksi adapteri, eli yksi DNA-fragmentti. Helmen pinnalla on kuitenkin useampia kiinnityskohtia, joihin monistumisvaiheessa templaatti-dnafragmentin perusteella monistetut juosteet kiinnittyvät. Jokainen DNA-fragmentti monistuu siis omassa helmessään. Kun jokainen fragmentti on kiinnittynyt omaan helmeensä, helmet lisätään seokseen, jossa DNA:n monistuminen tapahtuu. PCR-reaktio päättyy, kun helmien 16
.")
19 jokaisessa kiinnittymiskohdassa on juoste, ja helmien ympärillä oleva seos hajoaa. (Life Technologies Corporation 2013). Helmet pipetoidaan mikrosirulle, jossa on ionisensori ja yli miljoona mikroskooppista kaivoa, joihin yksittäiset helmet juosteineen asettuvat. Sekvensointilaite huuhtelee tätä sirua nukleotidi (Adeniini, Tymiini, Guaniini, Sytosiini) kerrallaan. Kun DNA-polymeraasi yhdistää nukleotidin templaatti-dna-juosteeseen, vapautuu vetyioni, joka aiheuttaa muutoksen ph:ssa. Kaivon alla oleva ionisensori aistii tämän muutoksen ph:ssa ja havaitsee, liitetäänkö kyseinen nukleotidi juosteeseen vai ei (Flushberg ym. 2010). Tämän biokemiallisen tiedon perusteella sekvensaattori rakentaa sekvenssit digitaaliseen muotoon mikrosirulle. Tätä toistetaan niin kauan kunnes sekvensointi on valmis (Life Technologies Corporation 2013) Datan klusterointi Sekvenssit analysoitiin QIIME (Quantitative insights Into Microbial Ecology) -ohjelmistolla (Caporaso ym. 2010), jonka avulla sekvenssit ryhmiteltiin operatiivisiksi taksonomisiksi yksiköiksi eli OTUiksi 97 prosentin samankaltaisuusasteella Usearch-algoritmia käyttäen (Edgar 2010). Analyysistä poistettiin huonolaatuiset ja liian lyhyet sekvenssit (katso Heino ym. 2015). Lajikertymät suhteutettiin pienimpään näytekokoon eli rarefoitiin bakteereilla ja sienillä 4104 sekvenssiin. Analyysissä tunnistettiin erilaista OTUa bakteeriyhteisöissä ja 3581 sieniyhteisöissä. Tilastollisista analyyseistä poistettiin ne OTUt, joita esiintyi vain 1 yksilö yhdellä paikalla (singletonit). Rarefoinnin ja sigletonien poistamisen jälkeen analyyseihin sisällytettiin 7290 (harva) ja 3798 (tiheä) bakteeri-otua sekä 1557 (harva) ja 1376 (tiheä) sieni-otua. OTUjen taksonomia määritettiin BLAST-työkalun avulla, joka vertaa määritettävää sekvenssiä NCBI-geenitietokantaan (The National Center of Biotechnology Information), ja antaa parhaan tuloksen perustuen sekvenssien samankaltaisuuteen (Altschul ym. 1990). Osa tutkimuspaikoista jouduttiin poistamaan analyyseistä, koska DNA-aineisto oli puutteellista: DNA:n monistuminen epäonnistui tai sekvenssejä oli liian vähän. Tiheiden lehtikarikepussien osalta neljän paikan aineisto ei ehtinyt mukaan analyyseihin. Tutkimuspaikkojen määrä (n) eri yhteisöanalyyseissä vaihteli seuraavasti: Harvojen pussien bakteerit (n=31), harvojen pussien sienet (n=32), tiheiden pussien bakteerit (n=26), tiheiden pussien sienet (n=24). 17

20 2.6 Tilastolliset analyysit Tilastolliset analyysit toteutettiin R-ohjelmistolla (R versio 3.2.0), paketeilla vegan (Oksanen ym. 2016), stats (R Core Team 2016) sekä labdsv (Roberts 2016). Yhteisöjen lajistovaihtelun ja paikkakohtaisen ryhmittymisen tarkasteluun käytin epämetristä moniulotteista pisteytystä eli NMDS-ordinaatiomenetelmää (engl. non-metric multidimensional scaling, paketti vegan). NMDS on monimuuttujamenetelmä, jossa pyritään kuvaamaan mahdollisimman vähäulotteisesti moniulotteista aineistoa. Epämetrisenä menetelmänä NMDS soveltuu hyvin aineistoille, jotka eivät ole normaalisti jakautuneita tai jatkuvia (McCune & Grace 2002). NMDS-ordinaation kuvaajassa pisteiden välinen välimatka kuvastaa yhteisöjen samankaltaisuuden tasoa, ja paikkapisteiden etäisyydet vastaavat mahdollisimman hyvin yhteisöjen välisiä laskennallisia etäisyyksiä. Etäisyysmittana tutkimuksessani käytin Bray-Curtisin samanlaisuusindeksiä, jota suositellaan käytettävän ekologisten yhteisöjen koostumuksen erojen mittaamiseen (McCune & Grace 2002). Analyysin stressiarvo kertoo, kuinka hyvin ordinaatioanalyysin tulos sopii alkuperäiseen aineistoon: mitä pienempi stressiarvo, sitä paremmin tulos kuvaa aineistoa. Stressiarvo kasvaa ulottuvuuksien vähentyessä, mutta toisaalta kuvaajan tulkitseminen muuttuu sitä vaikeammaksi, mitä moniulotteisempana se esitetään. Tässä aineistossa käytin kolmea ulottuvuutta, koska stressiarvo pieneni merkittävästi kaksiulotteisesta ratkaisusta ja analyysin tulkinta säilyi suhteellisen selkeänä. Ennen NMDSordinaatioanalyysiä lajistoaineistoon sovitettiin logaritmimuunnos log(x+1), missä X on havaitun taksonin yksilö/sekvenssimäärä. Muutoksen avulla dominoivien lajien vaikutusta analyyseihin pyrittiin tasoittamaan. Ympäristömuuttujat sovitettiin NMDS-ordinaatioon envfit-komennolla (paketti vegan), jossa ympäristötekijöiden ja ordinaation korrelaatioiden merkitsevyyttä testattiin permutaatiotestillä (1000 permutaatiota). Yhteisöjen eroavaisuuksia eri käsittelyiden välillä tarkasteltiin PERMANOVA-testillä (permutational analysis of variance; funktio Adonis paketissa vegan). Menetelmä on erityisesti ekologisiin aineistoihin soveltuva tilastollinen testi, jonka avulla voidaan tarkastella lajistokoostumuksien eroja tutkittavien ryhmien (tässä puroryhmät: luonnontilaiset, hiekoittuneet, kunnostetut) välillä (Anderson 2001). Analyysi perustuu permutaatioihin, eli matemaattiseen menetelmään, jossa testattavia arvoja järjestellään uudestaan suhteessa toisiinsa ja näin testataan, miten harvinainen havaittu tulos on. Tässä käytettiin 999 permutaatiota. 18
.")
21 Pohjaeläin- ja mikrobiyhteisöille laskettiin paikka- sekä käsittelykohtaiset diversiteettiindeksit, Shannonin diversiteetti-indeksi (lajimäärä) sekä Pieloun lajien tasaisuusindeksi (Pielou evenness) (Oksanen 2016). Käytin varianssianalyysiä sekä parittaista t-testiä (ANOVA ja pairwise t-test, paketti stats) ryhmien ja käsittelyiden välisten erojen tarkastelemiseen. Varianssianalyysi ja parittainen t-testi kertovat, onko ryhmien keskilukujen välillä tilastollisesti merkitseviä eroja (van Enden 2008). Analyysit tehtiin uoman ympäristömuuttujille, hajotukselle, pohjaeläinten ravinnonkäyttöryhmille sekä biodiversiteetti-indekseille. Pohjaeläinyhteisöille suoritettiin Indval-indikaattorilajianalyysi (paketti labdsv), jonka avulla pyrittiin tunnistamaan eri käsittelyluokan ilmentäjälajeja. Indikaattorilajianalyysi laskee käsittelyluokittelun pohjalta lajeille indikaattoriarvon käyttäen suhteellista yleisyyttä ja suhteellista keskimääräistä runsautta. Lisäksi se antaa luokan, jossa kullakin lajilla on suurin indikaattoriarvo sekä arvon ko. luokassa. Se myös laskee todennäköisyyden saada yhtä korkea indikaattoriarvo tietyssä määrässä toistoja. Täydellinen indikaattorilaji esiintyy kaikissa ko. luokan näytteissä eikä esiinny muissa luokissa (Dufrêne & Legendre 1997). Tutkin lehtikarikkeen hajoamisen suhdetta biologisiin ja ympäristömuuttujiin lineaariregression avulla. Sopivan mallin hakemiseen käytin askeltavaa regressioanalyysia. Lähdin rakentamaan mallia step-algoritmin avulla yhdessä lm-funktion (paketti stats) kanssa, joka rakentaa parhaan mallin perustuen AIC-arvoon (Akaike Information Criteria). Mallin rakentaminen tapahtui seuraavalla tavalla: 1) Ensiksi rakensin nollamallin (ei yhtään selittävää muuttujaa) ja kokonaismallit (kaikki muuttujat) erikseen uoman fysikaaliskemiallisille, habitaatti-, maankäyttö-, uoman ulottuvuus- ja biologisille muuttujille. Jos kokonaismallissa esiintyi muuttujien välistä korrelaatiota, osa muuttujista jätettiin pois mallista. Nollamallin pohjalta step-algoritmi lähtee rakentamaan parasta mallia askel askeleelta sekä eteenpäin- että takaisinvalinnalla suhteessa kokonaismalliin. Etenevässä valinnassa kerran mukaan otettu muuttuja ei voi enää pudota pois, ja vastaavasti takaisinvalinnassa kerran pudotettu muuttuja ei voi enää palata malliin (Holopainen & Pulkkinen 2002). Merkitsevimmät eli parhaaseen step-algoritmilla rakennettuun malliin sisällytetyt ympäristömuuttujat sisällytin analyysin seuraavaan vaiheeseen. 2) Edelleen rakensin nollamallin ja kokonaismallin merkitsevimmille muuttujille, joiden pohjalta stepalgoritmi rakensi lopullisen mallin edellä mainitulla tavalla. Testasin myös, lisääkö aikaisemmassa vaiheessa poistettujen ympäristömuuttujien lisääminen mallin selitysarvoaja luotettavuutta tarkastelemalla mallin selityskerrointa R 2, AIC-arvoa sekä p-arvoa. 19
22 Ympäristömuuttujien normaalijakautuneisuus tarkistettiin Shapiro-Wilk-testillä ja tehtiin logaritmi- tai neliöjuurimuunnos, jos se paransi muuttujan normaalijakautuneisuutta. Hajotusregressiomalleihin sisällytettiin 28 paikkaa (harvat lehtikarikepussit) ja 22 paikkaa (tiheät lehtikarikepussit), eli ne paikat, joista oli saatavilla onnistuneet biologiset yhteisöaineistot. 3. TULOKSET Luonnontilaisten purojen valuma-alueesta keskimäärin 5,5 prosenttia (vaihteluväli 0 13) oli ojitettu ja hienojakoisen aineksen osuus uoman pohjan peittävyydestä oli noin 20 prosenttia (vaihteluväli 10 33). Vastaavat luvut hiekoittuneille puroille olivat 20 (vaihteluväli 9 44) ja 50 (vaihteluväli 33 97) prosenttia sekä kunnostetuille puroille 25 (vaihteluväli 3 65) ja 40 (vaihteluväli ) prosenttia. Hienojakoisen aineksen määrä luonnontilaisissa puroissa oli merkittävästi pienempi kuin hiekoittuneissa ja kunnostetuissa puroissa (ANOVA, P < 0,05) (kuva 2). Vastaavasti raekoko oli luonnontilaisissa puroissa merkittävästi suurempi kuin hiekoittuneissa puroissa (parittainen t-testi, P < 0,001), ja kunnostetut purot erosivat lähes merkitsevästi vertailupuroista sekä hiekoittuneista puroista (parittainen t-testi, P < 0,1) (kuva 2). Kunnostettujen purojen vaihtelusta nähdään, että kunnostusten ansiosta hiekan määrä oli vähentynyt ja raekoko kasvanut osassa puroista, ja raekoko erosi lähes merkitsevästi vertailupuroista (parittainen t-testi, P < 0,1). Hiekan osalta ero ei ollut tilastollisesti merkitsevä (parittainen t-testi, P > 0,2). Sammalpeittävyys sekä puun määrä uomassa olivat merkitsevästi suurempia ja Pfankuch-luku merkitsevästi pienempi luonnontilaisissa puroissa (ANOVA, P < 0,05). Hiekan määrä korreloi negatiivisesti sammalpeittävyyden kanssa (-0,55, P < 0,005). Uoman hydrologis-morfologisesta muuttuneisuudesta kertova Pfankuch-luku korreloi positiivisesti hiekan määrän (0,74, P < 0,0001) ja negatiivisesti pohjan raekoon (-0,75, P < 0,0001) kanssa sekä sammalpeittävyyden (-0,72, P < 0,0001). Sammalpeittävyys korreloi positiivisesti pohjan raekoon kanssa (0,64, P < 0,0001). Veden väri korreloi negatiivisesti valuma-alueen mineraalimaan prosenttiosuuden kanssa (-0,71, P < 0,0001). Purot erosivat veden laadultaan, ojitusta lukuun ottamatta muulta maankäytöltään sekä maaperän ominaisuuksiltaan vain vähän toisistaan. Metsätyypissä (havumetsä, lehtimetsä, sekametsä, aukea eli matala kasvillisuus) oli jonkin verran vaihtelua. Tarkemmat purokohtaiset tiedot maankäytöstä ja mitatuista ympäristömuuttujista löytyvät liitteestä 1. 20
23 Kuva 3. Uoman pohjan keskimääräinen raekoko (Wenworthin asteikko) ja hiekan keskimääräinen peittävyys (%) uomasta eri käsittelyissä. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot Yhteisörakenne Mikrobiyhteisöjen taksonomia Noin 98 prosenttia bakteeriyhteisöjen BLAST-analysoiduista OTUista luokiteltiin vähintään pääjaksotasolle. Yhteensä eri taksonomiatasolle luokiteltuja eliöitä oli 946. Yleisimmät havaitut pääjaksot olivat Bacteroidetes sekä Proteobacteria (kuva 4). Harvojen lehtikarikepussien yhteisöissä myös ryhmän Firmicutes osuus oli huomattavan suuri. Muita ryhmiä olivat esimerkiksi Acidobacteria ja Actinobacteria. Bacteroidetes-ryhmän yleisin suku oli harvoissa pusseissa Bacteroides ja tiheissä pusseissa Flavobacterium. Ryhmässä Proteobacteria heimojen Comamonadaceae, Oxalobacteraceae ja Rhodocyclaceae määrittämättömät suvut olivat yleisimpiä. Sieniyhteisöjen BLAST-analysoiduista OTUista n. 40 prosenttia tunnistettiin vähintään pääjaksotasolle. Yhteensä eri taksonomiatasolle luokiteltuja eliöitä oli 478. Yleisin havaittu pääjakso oli Ascomycota, jonka osuus kaikista luokitelluista OTUista oli harvoissa lehtikarikepusseissa noin 85 % ja tiheissä noin 70 %. Toiseksi merkittävin ryhmä oli Basidiomycota, jonka osuus harvojen pussien yhteisöstä oli hieman yli 10 %, ja tiheiden pussien yhteisöistä noin 18 % (kuva 5). Tiheissä pusseissa merkittäväksi ryhmäksi nousi myös Blastocladiomycota, jonka osuus yli noin 10 % kaikista OTUista. Yleisin luokiteltu ryhmä oli suku Meliniomyces, jonka osuus Ascomyta-ryhmästä oli noin 80 %. Basidiomycota-ryhmän yleisin suku oli Cystolepiota ja Blastocladiomycota-ryhmän ainoa suku oli Blastocladiella. 21
24 HARVA TIHEÄ 100 % 100 % 90 % 90 % 80 % 80 % 70 % 70 % 60 % 60 % 50 % 50 % 40 % 40 % 30 % 30 % 20 % 20 % 10 % 10 % 0 % REF IMP REST 0 % REF IMP REST Bacteroidetes Proteobacteria Bacteroidetes Proteobacteria Firmicutes Muut Firmicutes Muut Kuva 4. Harvojen ja tiheiden lehtikarikepussien bakteeriyhteisöjen yleisimpien taksonomisten ryhmien osuus kaikista BLAST-määritetyistä taksoneista. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. HARVA TIHEÄ 100 % 100 % 90 % 90 % 80 % 80 % 70 % 70 % 60 % 60 % 50 % 50 % 40 % 40 % 30 % 30 % 20 % 20 % 10 % 10 % 0 % REF IMP REST 0 % REF IMP REST Ascomycota Basidiomycota Ascomycota Basidiomycota Blastocladiomycota Muut Blastocladiomycota Muut Kuva 5. Harvojen ja tiheiden lehtikarikepussien sieniyhteisöjen yleisimpien taksonomisten ryhmien osuus kaikista BLAST-määritetyistä taksoneista. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. 22
25 Osuus (%) Pohjaeläinten yhteisökoostumus Tutkimuksessa havaittiin yhteensä 89 pääasiassa lajitasolle määritettyä pohjaeläintaksonia (liite 2). Suurin lajimäärä oli pilkkojien ravinnonkäyttöryhmässä, jossa havaittiin 28 taksonia. Pilkkojien (P), laiduntajien (L) ja suodattajien (S) osuus koko yhteisöstä oli samaa suuruusluokkaa, eli vajaa kolmannes, kun taas petojen osuus oli pienempi, keskimäärin 10 prosenttia koko yhteisöstä (kuva 6). PLS-ravintokäyttöryhmien osuuksissa oli suuntaa antavia eroja käsittelyiden välillä, mutta erot eivät olleet tilastollisesti merkitseviä (ANOVA, P > 0,05). Pilkkojien määrä hiekoittuneissa puroissa oli alhaisempi kuin kunnostetuissa ja vertailupuroissa. Sen sijaan suodattajien määrä oli hiekoittuneissa puroissa suurempi kuin vertailupuroissa sekä kunnostetuissa puroissa, missä suodattajien osuus oli alhaisin. Laiduntajien osuus vaihteli myös jonkun verran käsittelyiden välillä; vertailupuroissa osuus oli alhaisin ja kunnostetuissa puroissa korkein. Petojen määrissä ei ollut huomattavia eroja Pilkkojat Laiduntajat Suodattajat Pedot REF IMP REST Kaikki Kuva 6. Pohjaeläinten ravinnonkäyttöryhmien yksilömäärien prosentuaalinen osuus koko yhteisöstä (keskiarvo ± keskivirhe). REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Pilkkojissa yleisimmät taksonit olivat Taeniopteryx nebulosa (24 %), Leuctra sp. (16 %) ja Potamophylax sp. (12 %). Laiduntajissa yleisimmät lajit olivat Baetis rhodani (58 %) ja Baetis niger (21 %). Suodattajissa yleisimmät taksonit olivat Simulidae (87 %) ja Plectrocnemia conspersa (7 %). Dicranota sp. (40 %), Isoperla sp. (18 %) ja Diura nanseni (11 %) olivat yleisimmät pedot. Lehtikarikepussien pohjaeläimistö koostui suurimmaksi osaksi pilkkojista (70 %), muiden ryhmien edustaessa noin 10 prosenttia pussien yhteisöistä. 23

26 Yleisimmät havaitut taksonit olivat Chironomidae, Limnephilidae sp., Nemoura sp. ja Taeniopteryx nebulosa. Indikaattorilajianalyysissa havaittiin viisi luonnontilaisia vertailupuroja edustavaa lajia, missä nämä lajit olivat suhteellisesti yleisempiä verrattuna hiekoittuneisiin ja kunnostettuihin puroihin (Taulukko 3): pilkkojalajit Taeniopteryx nebulosa ja Micrasema gelidum, laiduntajiin kuuluva Ameletus inopinatus, sekä pedot Diura nanseni ja Agabus sp. Hiekoittuneita puroja ilmensi huomattavan yleinen Capnopsis schilleri, ja kunnostusvaikutusta indikoi Potamophylax sp. (Taulukko 3). Taulukko 3. Pohjaeläinten indikaattorilajit sekä niiden esiintyminen ja suhteellinen yleisyys eri käsittelyissä. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Indikaattorilaji FFG* Käsittely Indikaattoriarvo p-arvo Esiintyminen Suhteellinen yleisyys** REF IMP REST REF IMP REST Micrasema gelidum SH REF 0,739 0,001 9/9 8/11 4/9 5,4 2,3 0,6 Taeniopteryx nebulosa SH REF 0,632 0,005 9/9 7/11 6/9 24,4 15,8 6,2 Ameletus inopinatus GRAZ REF 0,577 0,025 8/9 8/11 3/9 13,5 4,7 0,2 Diura nanseni PRED REF 0,569 0,016 7/9 7/11 5/9 24,7 7,0 3,0 Agabus sp. PRED REF 0,502 0,014 7/9 2/11 1/9 3,0 1,5 0,5 Capnopsis schilleri SH IMP 0,521 0,042 3/9 8/11 5/9 0,2 5,4 0,3 Potamophylax sp. SH REST 0,491 0,041 9/9 9/11 9/9 6,1 14,6 18,0 *Ravinnonkäyttöryhmä (Functinal Feeding Group) ** Suhteellinen yleisyys ravinnonkäyttöryhmässä EPT-taksonien (Ephemeroptera, Plecoptera ja Trichoptera) osuus oli jokseenkin alhaisempi hiekoittuneissa puroissa kuin muissa käsittelyissä. Sen sijaan CCS-sääskentoukkataksonien (Chironomidae, Ceratopogonidae ja Simuliidae) osuus hiekoittuneiden paikkojen yhteisöissä oli jokseenkin suurempi (kuva 7). Erot eivät olleet kuitenkaan tilastollisesti merkitseviä. Kuva 7. EPT- taksonien (Ephemeroptera, Plecoptera ja Trichoptera) ja CCS-taksonien (Chironomidae, Ceratopogonidae ja Simuliidae) yksilömäärien osuus koko pohjaeläinyhteisöstä eri käsittelyissä. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. 24
27 3.1.3 NMDS-yhteisöordinaatiot Harvojen lehtikarikepussien bakteeriyhteisöissä ei havaittu eroja yhteisökoostumuksessa eri puroryhmien suhteen. Bakteerien OTU-taksonit näyttävät jakautuvan ordinaation akseleiden 1 ja 2 suhteen kahteen ryhmään (kuva 8). Ordinaation akselin 1 ja 3 yhteisömuutos näyttää liittyvän habitaattiominaisuuksiin ja akselin 2 muutos veden fysikaalis-kemiallisiin ominaisuuksiin. Ordinaatioon sovitetuista ympäristömuuttujista vain muutama liittyi yhteisövaihteluun merkitsevästi (taulukko 4). Yhteisövaihtelu korreloi merkittävimmin akselilla 1 pohjoiskoordinaatin, akselilla 2 lämpötilan, uoman gradientin ja pohjoiskoordinaatin sekä akselilla 3 veden väriluvun, sammalpeittävyyden ja pohjan keskimääräisen raekoon suhteen (taulukko 4). Kuva 8. Harvojen lehtikarikepussien bakteerien OTU-taksonien ja tutkimuspaikkojen sijoittuminen NMDS-ordinaatioon (lopullinen stressi 0,031). ) Harmaat merkit (+) kuvaavat yhteisön eri taksoneita ja pisteet tutkimuspaikkoja. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Kuvaan sallittu ympäristömuuttujat, joiden p-arvo on korkeintaan (p.max) 0,2. 25
28 Harvojen lehtikarikepussien sieniyhteisöissä ei havaittu huomattavaa käsittelykohtaista ryhmittymistä (kuva 9). Luonnontilaisisten ja hiekoittuneiden purojen yhteisöjen parivertailussa havaittiin kuitenkin yhteisökoostumuksen merkitsevää erottumista (PERMANOVA, P < 0,05). OTU-taksonien yhteisörakenne näyttäisi vaihtelevan uoman fysikaalis-kemiallisten tekijöiden suhteen. Ordinaation akseleiden 1 ja 2 suhteen sovitetut ympäristömuuttujat näyttävät kuvaavan hyvin yhteisövaihtelun suuntaa, akselilla 3 yhteisövaihtelu ei ole suurta, ja näin tämän akselin suhteen sovitetut ympäristömuuttujat eivät kuvaa kovin hyvin yhteisövaihtelun suuntaa. Akseleiden kanssa merkittävimmin korreloivat veden väri, ph ja uoman leveys akselilla 1, valuma-alueen ojitusprosentti akselilla 2 sekä havumetsän osuus veden kiintoainepitoisuus, itäinen koordinaatti, pohjoinen koordinaatti sekä korkeus merenpinnasta (Alt) akselilla 3 (taulukko 4). Kuva 9. Harvojen lehtikarikepussien sienten OTU-taksonien ja tutkimuspaikkojen sijoittuminen NMDS-ordinaatioon (lopullinen stressi 0,1). Harmaat merkit (+) kuvaavat yhteisön eri taksoneita ja pisteet tutkimuspaikkoja. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Kuvaan sallittu ympäristömuuttujat, joiden p-arvo on korkeintaan (p.max) 0,2. 26
29 Taulukko 4. NMDS-yhteisöordinaatioihin sovitetut merkittävimmät ympäristömuuttujat ja korrelaatiot eri akseleiden suhteen. HARVAT BAKTEERIT TIHEÄT BAKTEERIT POHJAELÄIMET Muuttuja NMDS1 NMDS2 r2 Pr(>r) Muuttuja NMDS1 NMDS2 r2 Pr(>r) Lämpötila -0,138 0,990 0,404 0,001 *** Väri 0,999-0,044 0,542 0,001 *** Gradientti 0,363-0,932 0,205 0,044 * ph -0,977 0,215 0,421 0,002 ** Pohjoinen 0,781 0,625 0,199 0,045 * Leveys -0,821 0,571 0,296 0,007 ** ph -0,297-0,955 0,147 0,129 Ojitus -0,345 0,939 0,196 0,058. Pfankuch -0,859-0,511 0,118 0,174 Lämpötila 0,805-0,593 0,125 0,161 V.alueen koko -0,853-0,522 0,101 0,200 Muuttuja NMDS3 NMDS2 r2 Pr(>r) Muuttuja NMDS3 NMDS2 r2 Pr(>r) Lämpötila -0,072 0,997 0,346 0,003 ** Itä 0,993-0,115 0,221 0,029 * Väri -0,841 0,541 0,203 0,048 * Havumetsä -0,955 0,295 0,190 0,039 * Sammalpeittävyys -0,965 0,262 0,174 0,065. Kiintoaine 0,977 0,214 0,184 0,057. Rae -1,000-0,003 0,149 0,095. Ojitus -0,399 0,917 0,171 0,068. ph 0,760-0,650 0,163 0,101 Pohjoinen 1,000 0,018 0,157 0,082. V.alueen koko 0,970 0,243 0,111 0,185 Alt 0,999-0,051 0,146 0,091. Hiekka 0,999-0,043 0,111 0,198 V.alueen koko 0,971-0,239 0,135 0,122 Uoman varjostus -0,998 0,065 0,118 0,155 Muuttuja NMDS1 NMDS2 r2 Pr(>r) Muuttuja NMDS1 NMDS2 r2 Pr(>r) Lämpötila 0,577-0,817 0,812 0,001 *** Puun määrä -0,047-0,999 0,416 0,003 ** Väri 0,994-0,105 0,526 0,001 *** Ojitus -0,642 0,766 0,383 0,006 ** ph -0,983-0,186 0,464 0,004 ** Väri 0,625 0,780 0,300 0,023 * Leveys -0,881-0,472 0,406 0,007 ** Lämpötila -0,134 0,991 0,215 0,067. Ojitus -0,675-0,738 0,205 0,076. Leveys -0,981 0,192 0,224 0,069. Pfankuch 0,510 0,860 0,218 0,074. Muuttuja NMDS1 NMDS3 r2 Pr(>r) Muuttuja NMDS3 NMDS2 r2 Pr(>r) Väri 0,925-0,379 0,568 0,001 *** Alt 0,965-0,262 0,501 0,002 ** ph -0,952 0,307 0,475 0,004 ** Puun määrä 0,469-0,883 0,462 0,002 ** Leveys -0,997 0,077 0,328 0,017 * Sammalpeittävyys 0,857-0,515 0,342 0,014 * Lämpötila 1,000 0,010 0,300 0,020 * Ojitus -0,807 0,591 0,326 0,016 * Rae 0,451 0,892 0,290 0,030 * Pfankuch -0,834 0,552 0,273 0,038 * Hiekka -0,611-0,792 0,263 0,050 * Lämpötila 0,669 0,743 0,277 0,039 * Pfankuch 0,013-1,000 0,234 0,055. Väri 0,785 0,619 0,252 0,045 * Itä -0,467 0,884 0,200 0,067. Pohjoinen 0,937-0,350 0,234 0,063. Muuttuja NMDS1 NMDS2 r2 Pr(>r) Muuttuja NMDS3 NMDS2 r2 Pr(>r) Lämpötila 0,989-0,149 0,740 0,001 *** Sammalpeittävyys -0,581 0,814 0,456 0,001 *** Alt 0,164 0,987 0,428 0,002 ** Alt -0,377 0,926 0,459 0,001 *** Sammalpeittävyys 0,224 0,975 0,359 0,004 ** Ojitus 0,227-0,974 0,377 0,003 ** Ojitus 0,013-1,000 0,362 0,004 ** Pfankuch 0,936-0,352 0,304 0,015 * V.alueen koko 0,949 0,314 0,296 0,005 ** Hiekka 0,996-0,089 0,238 0,038 * Järvet 0,831 0,557 0,241 0,016 * Pohjoinen -0,760 0,650 0,238 0,038 * Väri 0,995 0,104 0,166 0,088. Puun määrä -0,579 0,816 0,211 0,046 * Raekoko -0,971 0,240 0,192 0,044 * HARVAT SIENET TIHEÄT SIENET POHJAELÄIMET Tiheiden lehtikarikepussien yhteisöissä oli nähtävissä jokseenkin käsittelykohtaista ryhmittymistä tutkimuspaikkojen suhteen, ja merkittävien ympäristömuuttujien joukossa on hiekoittumishäiriöstä kertovia muuttujia (Kuva 10). Luonnontilaisten purojen yhteisökoostumus erosi merkitsevästi hiekoittuneista puroista (PERMANOVA, P < 0,05). Akseleiden 1 suhteen merkittävimmin korreloivat lämpötila, veden väriluku, ph ja uoman leveys, akselin 2 suhteen valuma-alueen ojitusprosentti ja lämpötila sekä akselin 3 suhteen pohjan raekoko, hiekan peittävyys uomassa, Pfankuch-luku ja itäinen koordinaatti (taulukko 4). 27
30 Kuva 10. Tiheiden lehtikarikepussien bakteerien OTU-taksonien ja tutkimuspaikkojen sijoittuminen NMDS-ordinaatioon (lopullinen stressi 0,117). Harmaat merkit (+) kuvaavat yhteisön eri taksoneita ja pisteet tutkimuspaikkoja. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Kuvaan sallittu ympäristömuuttujat, joiden p-arvo on korkeintaan (p.max) 0,1. Tiheiden lehtikarikepussien sieniyhteisöissä ei havaittu tutkimuspaikkojen huomattavaa ryhmittymistä käsittelyiden suhteen (kuva 11). Yhteisövaihtelun kanssa merkittävimmin korreloivat uoman leveys, valuma-alueen ojitusprosentti ja veden väriluku akselilla 1, akselilla 2 puun määrä, valuma-alueen ojitusprosentti, veden väriluku, Pfankuch-luku ja lämpötila akselilla 2 sekä korkeus merenpinnasta (alt), sammalpeittävyys, valuma-alueen ojitusprosentti, sammalpeittävyys, Pfankuch-luku, lämpötila, veden väriluku ja pohjoiskoordinaatti akselilla 3 (taulukko 4). Luonnontilaiset vertailupaikkojen pohjaeläinyhteisöjen koostumus erosi merkittävästi hiekoittuneista ja kunnostetuista paikoista (PERMANOVA, P < 0,05). Myös lajistoordinaatiossa on havaittavissa käsittelykohtaista ryhmittymistä, etenkin akseleiden 2 ja 3 suhteen (kuva 12). Yhteisövaihtelun suhteen merkittävimmin korreloivat lämpötila, uoman valuma-alueen koko, valuma-alueen järvisyys sekä jokseenkin myös veden väriluku akselilla 1, valuma-alueen ojitusprosentti, sammalpeittävyys, korkeus merenpinnasta ja puun määrä uomassa akselilla 2, sekä Pfankuch-luku, pohjoiskoordinaatti, hiekan peittävyys ja pohjan raekoko akselilla 3 (taulukko 4). 28
31 Kuva 11. Tiheiden lehtikarikepussien sienten OTU-taksonien ja tutkimuspaikkojen sijoittuminen NMDS-ordinaatioon (lopullinen stressi 0,07). Harmaat merkit (+) kuvaavat yhteisön eri taksoneita ja pisteet tutkimuspaikkoja. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Kuvaan sallittu ympäristömuuttujat, joiden p-arvo on korkeintaan (p.max) 0,1. Kuva 12. Pohjaeläintaksonien ja tutkimuspaikkojen sijoittuminen NMDS-ordinaatioon (lopullinen stressi 0,117). Harmaat merkit (+) kuvaavat yhteisön eri taksoneita ja pisteet tutkimuspaikkoja. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. Kuvaan sallittu ympäristömuuttujat, joiden p-arvo on korkeintaan (p.max) 0,1. 29
32 3.1.4 Lajimäärä ja lajien tasaisuus Lajimäärässä ja lajien tasaisuudessa ei havaittu merkitseviä eroja käsittelyiden välillä mikrobiyhteisöissä. Bakteerien lajimäärä ja tasaisuus oli keskimäärin alempi hiekoittuneissa kuin kunnostetuissa ja vertailupuroissa, kun taas sienillä tilanne oli päinvastainen. Myöskään pohjaeläimillä ei havaittu merkitseviä eroja lajimäärässä tai lajien tasaisuudessa. Lajimäärä oli keskimäärin korkein luonnontilaisissa ja alhaisin kunnostetuissa puroissa. 3.2 Hajotus Mikrobien ja pohjaeläinten osuus lehtikarikkeen hajotuksesta arvioitiin jakamalla harvojen lehtikarikepussien massahävikin prosentuaalinen osuus tiheiden lehtikarikepussien vastaavalla luvulla. Aineiden liukenemisen ja mikrobihajotuksen osuus hajotuksesta oli noin 90 prosenttia (vaihteluväli ) ja siten pohjaeläinhajotuksen sekä fysikaalisen kulumisen noin 10 prosenttia. Keskimäärin tutkimusjakson aikana lehtikarikkeesta oli hajonnut 42 % (vaihteluväli 34 54) harvoissa pusseissa ja 38 % (vaihteluväli 36 40) tiheissä pusseissa. Hajotusnopeus oli keskimäärin 0,023/päivä harvoissa lehtikarikepusseissa (vaihteluväli 0,017 0,034) ja 0,020/päivä tiheissä lehtikarikepusseissa (vaihteluväli 0,019 0,021). Hajotusnopeus erosi merkitsevästi harvojen ja tiheiden pussien välillä (ANOVA, P < 0,0001), mutta ei käsittelyiden välillä (kuva 13). Kuva 13. Hajotusnopeus (k) eri käsittelyissä harvoissa ja tiheissä lehtikarikepusseissa. REF = luonnontilaiset, IMP = hiekoittuneet ja REST = kunnostetut purot. 30
33 Regressioanalyysin perusteella (taulukko 4) harvojen lehtikarikepussien hajotusnopeutta selitti eniten pohjaeläinten lajimäärä yhdessä uoman varjostuksen kanssa, jotka selittivät 45 % vaihtelusta. Vastaavasti tiheiden lehtikarikepussien hajotusta selittivät lämpötila, ojitusprosentti ja sekametsän osuus. Tiheän hajotuksen malliin sisällytin lisäksi sienitaksonien (OTU) tasaisuusindeksin sekä sammalpeittävyyden, sillä näiden muuttujien lisääminen paransi mallin selitysarvoa. Yhdessä nämä muuttujat selittivät 67 % vaihtelusta. Vertailemalla havaittuja ja mallin perusteella sovitettuja arvoja toisiinsa havaitaan, että mallin sovitus on melko hyvä. Selittämättä jäävä vaihtelu on kuitenkin melko suurta, joten myös muut tekijät kuin malliin sisällytetyt muuttujat vaikuttavat hajoamisnopeuteen (kuva 14). Taulukko 4. Yhteenveto hajotusnopeuden (k) lineaariregressiomalleista: selittävien biologisten ja ympäristömuuttujien suhde hajotusnopeuteen. Vastemuuttuja R 2 P-arvo AIC Selittävä muuttuja Selityskerroin P-arvo Hajotus (HARVA) 0,45 <0, ,80 Pohjaeläinten lajimäärä 0, ,002 Uoman varjostus -0, ,015 Lämpötila 0, ,008 Sienitaksonien (OTU) tasaisuus -0, ,033 Hajotus(Tiheä) 0,67 0, ,40 Ojitusprosentti 0, ,003 Sekametsän osuus 0, ,011 Sammalpeittävyys 0, ,030 a) b) Kuva 14. Lineaariregression avulla mallinnettujen hajotusnopeusarvojen (k) suhde havaittuihin arvoihin a) harvoissa lehtikarikepusseissa b) tiheissä lehtikarikepusseissa. 31
34 4. POHDINTA Maankäytöstä aiheutuva lisääntynyt sedimentaatio, kuten hiekoittuminen, yksipuolistaa jokiuoman rakennetta, kun jokiuomaan kasaantuva hiekka peittää allensa karkeamman aineksen, vähentää uoman syvyysvaihtelua ja siten yksipuolistaa uoman virtausoloja (Wood & Armitage 1997, Kemp 2011). Myös tässä tutkimuksessa havaittiin metsäojitusten kautta lisääntyneen hiekoittumisen vaikuttaneen merkittävästi purouoman pohjan laatuun: hienojakoisen aineksen osuus uoman peittävyydestä oli kasvanut ja uoman pohjan keskimääräinen raekoko pienentynyt. Tutkimuksen perusteella osassa puroista kunnostuksilla oli onnistuttu poistamaan hiekkaa uomasta ja lisäämään uoman pohjan rakenteellista monimuotoisuutta. Näissä puroissa hiekan määrä oli oletettavasti vähentynyt, sillä niiden uoman pohjamateriaalin keskimääräinen raekoko oli suurempi kuin kunnostamattomissa puroissa. Kunnostettujen purojen joukon sisällä oli kuitenkin suurta vaihtelua: osassa kunnostettuja puroja kunnostuksilla ei oltu onnistuttu vähentämään hiekan peittävyyttä eikä näin ollen monipuolistamaan uoman rakennetta. Erot pohjan raekoossa eivät olleet tämän takia tilastollisesti merkitseviä kunnostettujen ja kunnostamattomien puroryhmien välillä. Yksi uoman monimuotoisuudesta ja luonnontilaisuudesta kertova merkki on uoman pohjan sammalpeittävyys, joka oli kunnostetuissa puroissa vielä hiekoittuneiden purojen tasolla. Sammalet ovat tärkeä osa puroekosysteemin ekologista hyvinvointia, sillä ne tarjoavat habitaatteja pohjaeläimille, lisäävät uoman rakenteellista monimuotoisuutta ja pidättävät hyvin orgaanista ainesta, ja näin vaikuttavat koko puroyhteisön rakenteeseen ja toimintaan (Vuori & Joensuu 1996). Sammalyhteisöjen on todettu elpyvän hitaasti kunnostuksen aiheuttamasta häiriöstä (Muotka & Laasonen 2002), mitä havaintoa myös tämän tutkimuksen tulokset tukevat. Mikrobiyhteisöt koostuivat tavanomaisesti virtavesissä tavattavista ryhmistä (Zegling 2015, Kuehn 2016). Suuri osa sienten OTU-taksoneista kuului tunnistamattomaan ryhmään, kuten aikaisemmissakin tutkimuksissa on huomattu (Bärlocher 2016), mikä kertoo siitä, että virtavesien sienilajisto on vielä puutteellisesti tunnettu. Tutkimuksessa havaittiin tuhansia eri taksonomisiksi yksiköiksi (OTU) määriteltyjä mikrobeja, mikä tekee mikrobiyhteisöjen ekologisten vasteiden tulkinnan jokseenkin haasteelliseksi (Bärlocher 2016). Mikrobiyhteisöjen koostumus ei pääosin eronnut eri käsittelyiden välillä, kuten oletettiin. Kuitenkin luonnontilaisten purojen harvojen lehtikarikepussien sieniyhteisöt ja tiheiden lehtikarikepussien bakteeriyhteisöt erosivat merkittävästi hiekoittuneiden purojen yhteisöistä PERMANOVA-analyysin perusteella. Hiekoittuminen on siten jokseenkin 32
35 vaikuttanut myös mikrobiyhteisöjen rakenteeseen. Mitatuista ympäristömuuttujista mikrobiyhteisöjen koostumusta näyttivät selittävän eniten uoman vesikemialliset tekijät, kuten lämpötila, ph ja veden väriluku, jotka olivat useimmiten yhteisövaihtelua merkittävimmin selittävien ympäristötekijöiden joukossa. Tämä tukee Heinon ja kollegoiden tutkimusta (2014), jotka havaitsivat vesikemiallisten muuttujien selittävän eniten mikrobiyhteisöjen vaihtelua ja uoman habitaattimuuttujien olevan selvästi vähemmän tärkeitä yhteisöjen muodostumisessa (Heino ym. 2014). Myös tässä tutkimuksessa uoman habitaatin laadusta kertovat muuttujat näyttäisivät selittävän jonkin verran mikrobien yhteisövaihtelua. Lisäksi maantieteelliset tekijät kuten valuma-alueen koko, maantieteellinen sijainti sekä korkeus merenpinnasta korreloivat yhteisökoostumuksen kanssa. Nämä havainnot tukevat aikaisempia tutkimuksia (Heino ym. 2014, Bergfur & Sundberg 2014). Habitaatin laatuun ja uoman kokoon liittyvien tekijöiden tiedetään määrittelevän pohjaeläinyhteisöjen koostumusta (mm. Jones ym. 2012, Richards ym. 1997, Heino ym. 2004). Myös tässä tutkimuksessa pohjaeläinten yhteisökoostumusta näyttivät selittävän eniten uoman habitaatin laatua kuvaavat muuttujat, kuten sammalpeittävyys, pohjan raekoko ja puun määrä uomassa, sekä valuma-alueen ominaisuuksiin liittyvät tekijät, kuten valumaalueen koko ja järvisyys. Tutkimuksen perusteella luonnontilaiset purot erosivat pohjaeläinyhteisökoostumukseltaan hiekoittuneista ja kunnostetuista puroista. Hiekoittumiskuormituksella on siis ollut huomattava vaikutus pohjaeläinyhteisöjen rakenteeseen, eivätkä kunnostustoimet ole edesauttaneet yhteisöjen palautumista luonnontilaisten purojen tasolle. Hiekoittumisen seurauksena purouoman hienojakoisen aineksen prosentuaalinen osuus pohjan alasta kasvaa, kun karkeampi aines ja kivien välitilat peittyvät hiekalla, ja habitaatti yksipuolistuu (Wood & Armitage 1997, Kemp ym. 2011, Jones ym. 2012). Hiekoittumisesta hyötyvät sellaiset lajit, jotka suosivat hienojakoista sedimenttiä elinympäristönään. Esimerkiksi hiekoittuneilla paikoilla suhteellisen yleisen ja indikaattorilajiksi luokitellun Capnopsis schillerin on havaittu suosivan hienojakoista pohjamateriaalia (Lillehammer 1975). Myös surviaissääskien ja muiden sääskiheimojen toukkien on havaittu hyötyvän hiekoittumisesta, sillä ne ovat suosivat hienojakoista pohjaa habitaattinaan (Richards ym. 1997, Sutherland ym. 2012). Sen sijaan herkkien EPT-lajien on havaittu taantuvan hiekoittumisen seurauksena (Sutherland ym. 2012, Vuori & Joensuu 1996, Martel ym. 2007). EPT-taksonien pohjaeläimet ovat pitkäikäisiä ja iso osa niistä suosii karkeampaa pohjamateriaalia habitaattinaan (Richards ym. 1997), joten ne ovat erityisen 33
36 herkkiä pohjan rakenteen muutoksille. Tässä tutkimuksessa EPT-taksonien suhteellisessa osuudessa pohjaeläinlajistossa oli havaittavissa laskua hiekoittumisen seurauksena, sen sijaan CCS-ryhmän sääskitoukkien osuudessa koko pohjaeläinyhteisöstä oli havaittavissa nousua hiekoittumisen suhteen. Luonnontilaisten purojen kaikki viisi indikaattorilajia kuuluvat EPT-ryhmään ja mukana oli sekä pilkkoja-, laiduntaja- että petolajeja, luultavasti uoman elinympäristö- ja yhteisömuutosta indikoivia häiriöherkkiä lajeja. Esimerkiksi Ameletus inopinatuksen on havaittu olevan herkkä sedimentaatiokuormitukselle (Tolkkinen ym. 2016). Nämä havainnot viittaavat siihen, että ympäristömuutokselle herkkien lajien tiheydet vähenevät, kun taas häiriötä hyvin sietävät generalistilajit yleistyvät hiekoittumisen seurauksena (Sutherland ym. 2012, Martel ym. 2007). Pohjaeläinten yksilömääriin perustuvissa ravinnonkäyttöryhmien osuuksissa havaittiin suuntaa antavia eroja purokäsittelyiden välillä, joskin havaitut erot jäivät vaille tilastollista tukea. Havaitut erot yhteisöissä voivat kuitenkin olla ekologisesti merkityksellisiä, ja ovat seurausta luultavasti hiekoittumiskuormituksen aiheuttamasta häiriöstä purouoman pohjan rakenteessa ja saatavilla olevan ravinnon laadussa (Jones ym. 2012). Suodattajat olivat yleisin ravinnonkäyttöryhmä hiekoittuneissa puroissa, mikä johtui mäkäräntoukkien Simuliidae yleistymisestä, sillä yli 90 % suodattajista oli mäkäriä. Myös Vuori ja Joensuu (1996) havaitsivat mäkäräntoukkien lisääntyneen metsäojitusten hiekoittamissa uomissa. Mäkäräntoukat sietävät melko hyvin häiriötä, ja ne luultavasti hyötyvät kiintoainekuormituksesta, kun veden mukana kulkeutuvan hienojakoisen orgaanisen aineksen määrää lisääntyy (Jones ym. 2012, Wallace & Webster 1996, Vuori & Joensuu 1996). Pohjan lisääntynyt epävakaus saattaa aiheuttaa negatiivisia muutoksia laiduntajien ja pilkkojien ravinnon saatavuudessa. Pohjan epävakaus haittaa vakaan leväbiofilmin muodostumista, josta laiduntajat ovat riippuvaisia (Jones ym. 2012, Wallace & Webster 1996), ja suspensoituneen kiintoaineen takia lisääntynyt veden sameus myös haittaa levien kasvua. Pilkkojien ravintona käyttämä lehtikarike hautautuu helposti liikkuvan sedimentin alle (Wallace & Webster 1996) tai kulkeutuu virran mukana alavirtaan (Muotka & Laasonen 2002), mikä voi vaikuttaa pilkkojien käytettävissä olevan ravinnon saatavuuteen ja pilkkojien yleisyyteen pohjaeläinyhteisössä. Osassa puroista kunnostustoimet olivat jokseenkin hyödyttäneet pilkkojayhteisöjä, johtuen luultavasti lehtien paremmasta pidättäytymisestä uomassa pilkkojalajien hyödynnettävissä (Lepori ym. 2005a, Muotka & Laasonen 2002, Flores ym. 2011). Laiduntajien osuus kunnostetuissa puroissa oli suurin ja luonnontilaisissa vertailupuroissa pienin. Tämä voi olla seurausta siitä, että kunnostettuihin 34
37 puroihin lisätty puuaines on toiminut kasvualustana pohjaleville (Wallace ym. 2015), joita laiduntajat hyödyntävät ravintonaan. Luonnontilaisissa puroissa vahva törmän kasvillisuus varjostaa puroja, jolloin valon määrä vähenee ja levien fotosynteesitehokkuus heikkenee (Hill 1996). Vahvan varjostuksen takia luonnontilaisten purojen leväbiotuotanto on rajoittunutta (Heino ym. 2004), minkä seurauksena laiduntajien osuus luonnontilaisissa yhteisöissä on luultavasti pienempi kuin hiekoittuneissa ja kunnostetuissa puroissa, joissa uoman varjostus on vähäisempää törmän muuttuneen kasvillisuuden takia. Kuitenkin havaitut suuntaa-antavat erot voivat johtua myös näytteenottoon liittyvästä satunnaisuudesta, sillä yhteisökoostumus voi vaihdella merkittävästi myös yksittäisen puron sisällä riippuen näytteenottopaikasta (Heino ym. 2004), joten tilastollisen tuen puutteen takia hiekoittumisen tai kunnostusten vaikutuksista pohjaeläinten toiminnalliseen ravinnonkäyttöryhmä-koostumukseen ei voida vetää luotettavia johtopäätöksiä. Kolmen viikon tutkimusajanjaksolla mikrobien osuus lehtikarikkeen hajotuksesta oli huomattavasti suurempi kuin pohjaeläinten. Myös Bergfur kollegoineen (2007) havaitsi mikrobien vastaavan suuresta osasta lehtikarikkeen alkuvaiheen hajotusta (Bergfur ym. 2007). Mikäli tutkimusta olisi jatkettu pidempään, olisi pohjaeläinten osuus hajotuksesta luultavasti kasvanut, sillä pohjaeläinten on havaittu vastaavan suurelta osin lehtikarikkeen hajotuksesta (esim. Hieber & Gessner 2002). Lehtikarikkeen hajoamisnopeus oli tavanomaisella hyvästä ekosysteemin tilasta kertovalla tasolla (Gessner & Chauvet 2002), eikä tutkimusten tulosten perusteella hiekoittumiskuormituksella tai purokunnostuksilla havaittu olevan merkittäviä vaikutuksia ekosysteemin lehtikarikkeen hajoamiseen. Hiekoittumisella on aikaisemmin havaittu ristiriitaisia tai olemattomia vaikutuksia lehtikarikkeen hajoamisnopeuteen (McKie & Malmqvist 2009, Matthaei ym. 2010, Sanpera- Calbet ym. 2012), kuin myös biodiversiteettiin (Heino ym. 2009, McKie & Malmqvist 2009), mikä kertoo siitä, että virtavesiekosysteemin vasteet hiekoittumishäiriölle voivat vaihdella riippuen häiriön voimakkuudesta ja ekosysteemin ominaisuuksista. Maltillisen häiriön seurauksena ekosysteemin eliöyhteisöjen koostumuksessa voidaan nähdä muutoksia, ja häiriön voimistuessa ekosysteemin toiminta, kuten orgaanisen aineen hajotusprosessi, usein heikkenee (Feld 2011). Tulosten perusteella voidaan sanoa, että hiekoittumiskuormitus on aiheuttanut lieviä muutoksia hajottajayhteisöissä, mutta ekosysteemin ekologinen toimintakyky ei ole kuitenkaan heikentynyt eikä biologinen monimuotoisuus menetetty hajotustoiminnan suhteen. 35
38 Lehtikarikkeen hajoamisnopeuteen vaikuttivat eniten pohjaeläinten lajimäärä (positiivinen) sekä uoman varjostus (negatiivinen) harvoissa pusseissa. Lajirikkauden ja hajotuksen yhteyttä on tutkittu paljon (Gessner ym. 2010), ja pohjaeläinten, etenkin pilkkojien (Jonsson ym. 2001, Dangles & Malmqvist 2004), lajirikkauden on havaittu nopeuttavan hajotusprosessia (mm. Webster & Benfield 1986, Lepori ym. 2005a, Graca 2001). Uoman varjostuksen negatiivinen vaikutus perustuu luultavasti siihen, että varjostus alentaa veden lämpötilaa ja vähentää saatavilla olevan valon määrää, ja sitä kautta hidastaa hajoamisnopeutta vaikuttamalla mikrobien metaboliseen aktiivisuuteen (Webster & Benfield 1986). Tiheissä lehtikarikepusseissa hajoamisnopeuteen vaikuttivat positiivisesti lämpötila, sekametsän osuus, ojitusprosentti ja sammalpeittävyys, sekä negatiivisesti sienitaksonien (OTU) tasaisuus. Metsätyyppi määrittelee uomaan ajautuvan lehtikarikkeen määrää ja laatua. Sekametsästä ajautuu ravintorikasta lehtikariketta uomaan, joten monipuolisen ravintoresurssin ylläpitämä hajottajayhteisö on luultavasti monipuolisempi kuin havumetsissä, ja siten lehtikarikkeen hajoaminen nopeampaa (McKie & Malqvist 2009, Yoshimura 2012). Sammalpeittävyys lisää uoman pohjan rakenteen monipuolisuutta ja lehtikarikkeen pidättäytymistä uomassa (Vuori & Joensuu 1996, Muotka & Laasonen 2002), ja siten vaikuttaa hajottajien ravintoresurssien määrään. Ojitusten aiheuttama kuormitus luultavasti nopeuttaa lehtikarikkeen hajoamista fyysisen kulumisen lisääntymisen kautta, kun veden mukana kulkeutuva hiekka kuluttaa lehtimateriaalia (Matthaei ym. 2010). Sieniyhteisöjen tasaisuuden on myös aikaisemmissa tutkimuksissa havaittu alentavan hajotusnopeutta (Tolkkinen ym. 2013), mikä kertoo siitä että sienilajien dominanssisuhteet vaikuttavat lehtikarikkeen hajoamisnopeuteen. Mitä enemmän hajottavia lajeja, sitä suurempi on kilpailu resurssista, ja eri lajit voivat toiminnallaan heikentää toistensa toimintaa esimerkiksi erittämällä kilpailijoiden metaboliaa heikentäviä yhdisteitä, ja siten hidastaa hajoamisprosessia (Costantini & Rossi 2010). Yleensä mikrobihajotus saturoituu, kun muutama laji dominoi hajotusprosessia (Gessner ym. 2010). Purokunnostukset olivat jokseenkin parantaneet uoman pohjan laatua osassa puroja, mutta näyttö kunnostusten onnistumisesta jäi vaille tilastollista tukea. Kunnostusten biologiset vasteet olivat heikot. Vaihtelu kunnostettujen purojen joukon sisällä oli suurta, mikä kertoneekin siitä, että osassa puroista kunnostukset olivat onnistuneet paremmin kuin toisissa. Tähän voivat olla syynä monet eri tekijät. Ensinnäkin, kunnostustoimet eri puroissa oli toteutettu osittain eri menetelmin ja eri mittakaavassa: osassa puroissa oli tehty vain pienemmän mittakaavan törmä- ja uomakunnostuksia, kun taas osassa paikoissa oli tehty 36
39 myös valuma-aluekunnostuksia (Yrjänä ym. 2011). Pelkät pienemmän mittakaavan törmäja uomakunnostukset voivat olla mitättömiä, mikäli suuremman mittakaavan prosessia, tässä tapauksessa liiallista hienojakoisen aineiden kulkeutumista uomaan, ei saada hallittua (Roni ym. 2008, Feld ym. 2011). Siksi valuma-alueen kunnostustoimet ovat erityisen tärkeitä hiekoittumisen ja sen ekologisten haittojen vähentämiseksi. Kuitenkin uoman sisäiset kunnostustoimet ovat myös tärkeitä, sillä ne monipuolistavat uoman pohjan rakennetta ja virtausoloja, ja sysäävät uoman luonnonmukaistamisprosessin käyntiin (Yrjänä ym. 2011). Luonnolliset veden kulkuun ja aineiden kulkeutumiseen liittyvät hydrogeomorfologiset prosessit muokkaavat ja ylläpitävät habitaatteja biologisille yhteisöille, joten näiden prosessien luonnonmukaistaminen sekä valuma-alue- että uomakunnostusten avulla on tarkoituksenmukaisempaa kuin pelkästään uoman pohjan rakenteen monipuolistaminen (Marttila ym. 2015). Toiseksi, kunnostuksista kulunut aika voi olla liian lyhyt: tutkimus tehtiin 3-9 vuotta kunnostusten jälkeen, eivätkä yhteisöt ja prosessit ole välttämättä ehtineet palautua häiriöstä. Muotka ym. (2002) havaitsivat merkittäviä eroja sammalpeittävyydessä ja pohjaeläinyhteisöissä riippuen siitä, kuinka pitkä aika purokunnostuksista oli kulunut. Esimerkiksi sammalyhteisöt vaativat 6-8 vuotta elpymiseen (Muotka ym. 2002). Etenkin biologisten yhteisöjen toipuminen voi olla hidasta, yhteisöjen sekä uoman ja valuma-alueen rakenteen palautuminen voi kestää jopa vuosikymmeniä (Wood & Armitage 1997, Vuori & Joensuu 1996, Leps ym. 2016, Lake ym. 2007). Kuitenkin biologiset vasteet ovat olleet myös pitkäaikaistutkimuksissa heikkoja (mm. Leps ym. 2016, Louhi ym. 2011). Kolmanneksi, biologisten yhteisöjen elpymiseen voivat vaikuttaa muut tekijät, kuten kolonisaatioprosessit ja biologisiin vuorovaikutussuhteisiin liittyvät tekijät (Lake ym. 2007, Sundermann ym. 2011). Kunnostusten heikot biologiset vasteet voivat olla seurausta esimerkiksi siitä, että puroyhteisöjä dominoivat edelleen ennen kunnostusta muodostuneet tolerantit yhteisöt, joten herkkien lajien palautumiselle ei ole yksinkertaisesti tilaa yhteisössä (Leps ym. 2016), tai niiden levittäytyminen takaisin purojaksolle on rajoittunut tai kokonaan estynyt (Lorenz & Feld 2013). Myös muissa tutkimuksissa havaitut biologiset vasteet ovat jääneet usein heikoiksi, vaikka useissa tutkimuksissa kunnostusten on todettu lisäävän uoman habitaattien monimuotoisuutta ja parantavan uoman fyysistä rakennetta kohti luonnontilaisempaa jokiuomaa (Haase ym. 2013, Leps ym. 2016, Feld ym. 2011, Louhi ym. 2011, Palmer ym. 2010, Jähning ym. 2009, Lepori ym. 2005a,b, mutta katso Miller ym. 2010). Tämä voi kertoa siitä, että vaikka muutos biologisissa yhteisöissä aiheutuisikin ihmisperäisestä häiriöstä, 37
40 esimerkiksi hiekoittumisesta, todellisuudessa biologisten yhteisöjen muodostumiseen vaikuttavat yhtä aikaa monet prosessit ja tekijät, joiden vuorovaikutuksia emme vielä täysin ymmärrä (mm. Palmer ym. 2010), ja siten yhteisöt eivät palaudu suoraviivaisesti häiriöstä. Toisaalta puroekosysteemeille on ominaista ympäristötekijöiden suuri ajallinen ja paikallinen vaihtelu (mm. Marttila ym. 2015), jolloin myös biologisissa yhteisöissä tapahtuu jatkuvia muutoksia. Siten tutkimusajankohdalla voi olla myös merkitystä siihen, millaisia vasteita yhteisöissä näemme, ja ympäristönvaihtelun satunnaisuuden takia voidaan helposti tehdä virhepäätelmiä. Tämä luonnollinen vaihtelu vaikeuttaa myös ihmisperäisten häiriöiden vasteiden tunnistamista (Underwood 1994). Tässä tutkimuksessa käytettiin kontrolli-painevertailuasetelmaa, jossa luonnontilaisia, kunnostettuja ja hiekoittuneita puroja verrattiin toisiinsa. Yksittäisten puroekosysteemien välillä esiintyvää luonnollista vaihtelua on vaikea hallita vertailemalla yksittäisistä puroista koostuvia joukkoja toisiinsa, jolloin aineiston tilastollinen voima heikentyy. Optimaalinen tilanne olisi, jos biologisia yhteisöjä olisi tutkittu ennen purokunnostuksia (ennen-jälkeen-kontrolli-paine-asetelma (BACI), Stewart- Oaten 1986). Tällöin yhteisövaihtelua voitaisiin vertailla sekä yksittäisen puron sisällä että purojoukoista koostuvien käsittelyiden välillä, jolloin luonnollisen vaihtelun tunnistaminen ja siten todellisten yhteisövasteiden havaitseminen olisi helpompaa (Underwood 1994). Todellisen yhteisömuutoksen havaitsemiseksi olisikin tärkeää tehdä pitkän aikavälin seurantatutkimuksia, jolloin ympäristön satunnaisvaihtelusta aiheutuva yhteisöjen luonnollinen ajallinen vaihtelu voidaan paremmin tunnistaa (Marttila ym. 2015, Huttunen ym. 2012). Tutkimukseen sisältyy aina epävarmuustekijöitä, jotka ovat voineet vaikuttaa tuloksiin. Tutkimusaineiston keräämiseen ja käsittelyyn osallistui monia eri henkilöitä, mikä voi lisätä mittausvirhettä aineistossa. Lehtikarikkeen hajotusprosessia tutkittiin ankkuroimalla lehtikarike rakennustiilien avulla uoman pohjaan. Uomaan sijoitettu tiili on voinut muuttaa paikallisia virtausolosuhteita, mikä voi vaikuttaa lehtikarikkeen hajoamiseen ja hajottajayhteisöjen muodostumiseen vaikuttaviin ympäristöolosuhteisiin (Young ym. 2008). Luonnossa uomaan päätyvä lehtikarike voi ajautua virran mukana, ja päätyy usein hitaamman virtauksen paikkoihin, joihin voi myös muodostua karikepatoja (Webster ym. 1999), tai pidättäytyy kivien, sammalien ja puumateriaalin ansiosta kovemman virtauksen paikoissa (Lepori ym. 2005a, Muotka & Laasonen 2002). Tämän huomioimiseksi lehtikarikepussit pyrittiin sijoittamaan uomaan ekologisesti mielekkäisiin paikkoihin, joihin lehtikarike myös luonnollisesti kerääntyisi (Boulton & Boon 1991). Vertailun 38
41 standardoimiseksi lehtikarikepussit pyrittiin sijoittamaan mahdollisimman samankaltaisiin paikkoihin eri uomissa, sillä habitaatti vaikuttaa hajoamisnopeuteen: esimerkiksi koskipaikoilla lehtikarikkeen hajoaminen on nopeampaa kuin suvantopaikoilla (Young ym. 2008). Jatkotutkimuksissa lehtikarikepussien sijoituspaikoilta kannattaisi tehdä pohjan laadun arviointi, mikä saattaisi parantaa tausta-aineiston selitysvoimaa, sillä habitaatin pohjan koostumus ja virtausolosuhteet vaikuttavat pohjaeläinten esiintymiseen ja siten siihen, mitkä pohjaeläimet lehtikarikepusseja kolonisoivat (Webster & Benfield 1986). Heikkojen vasteidensa takia on aiheellista pohtia, ovatko hajottajayhteisöt mielekkäitä organismeja hiekoittumisen aiheuttaman häiriön ja purokunnostusten ekologisen onnistumisen tutkimiseen. Etenkään vaikutukset mikrobiyhteisöihin eivät ole selkeitä ja huomattavia, koska mikrobiyhteisöihin ja niiden toimintaan vaikuttavat lähinnä veden laatuun liittyvät tekijät (Heino ym. 2014, Zegling 2015, Kuehn 2016). Siten hajotusprosessissakaan ei välttämättä havaita huomattavaa heikentymistä hiekoittumisen seurauksena, etenkään alle kuukauden tutkimusjaksolla, sillä hajotuksen alkuvaiheessa mikrobien osuus lehtikarikkeen kokonaishajotuksesta on usein huomattavampi kuin pohjaeläinten (Bergfur 2007). Sen sijaan myöhäisemmässä vaiheessa, kun mikrobit ovat muokanneet lehtikarikkeen koostumusta pohjaeläimille paremmin hyödynnettävään muotoon, pohjaeläinten merkitys hajotuksesta on suurempi (Webster & Benfield 1986, Hieber & Gessner 2002). Siten tutkimusajan pidentäminen voisi paljastaa vasteita myös hajotusprosessissa, koska pohjaeläinyhteisöissä havaittiin vasteita hiekoittumiselle. Sen sijaan kunnostuksilla ei havaittu vaikutuksia pohjaeläinyhteisöissä, mikä tukee aikaisempia havaintoja. Esimerkiksi Jähning kollegoineen (2009) ei havainnut kunnostusvasteita pohjaeläinyhteisöissä, mutta sen sijaan rannan tulvavyöhykkeen kasvillisuudessa havaittiin merkittäviä muutoksia. Niinpä tutkimuksen keskittäminen muihin organismeihin ja prosesseihin voisi paljastaa merkittävämpiä biologisia vaikutuksia etenkin purokunnostusten suhteen. 39
42 5. JOHTOPÄÄTÖKSET Tämän tutkimukset perusteella metsätaloustoimien takia lisääntynyt hiekkakuormitus oli yksipuolistanut purouoman pohjan rakennetta, sillä hiekan peittävyys pohjasta oli suurempi ja keskimääräinen raekoko pienempi metsätalousalueiden puroissa verrattuna luonnontilaisiin puroihin. Hiekoittumisen vähentämiseen pyrkivät purokunnostukset olivat onnistuneet vähentämään hiekkaa ja monipuolistamaan uoman pohjan rakennetta osassa kunnostetuista puroista. Kunnostettujen purojen joukon sisällä oli kuitenkin suurta vaihtelua: osassa kunnostettuja puroja kunnostukset eivät olleet onnistuneet vähentämään hiekan määrää eikä näin ollen monipuolistamaan uoman rakennetta, luultavasti erilaisten kunnostusmenetelmien takia. Luonnontilaisten purojen pohjaeläinyhteisökoostumus erosi merkittävästi hiekoittuneista ja kunnostetuista puroista, eli hiekoittuminen oli merkittävästi muuttanut pohjaeläinyhteisöjen rakennetta. Sen sijaan lehtikarikepussien mikrobiyhteisöissä havaittiin olemattomia tai heikkoja vasteita hiekoittumiselle, joten hiekoittuminen oli vaikuttanut heikosti mikrobiyhteisöjen koostumukseen. Lehtikarikkeen hajoamisprosessissa ei havaittu heikentymistä hiekoittumisen seurauksena. Biologisten yhteisöjen vasteet purokunnostuksille olivat olemattomia, mikä luultavimmin aiheutui riittämättömistä kunnostustoimenpiteistä tai liian lyhyestä vasteajasta kunnostusten jälkeen, ja hiekan aiheuttama häiriö uomassa jatkuu. KIITOKSET Ensiksi haluan kiittää työni ohjaajia dos. Jukka Aroviitaa, FM Jarno Turusta ja Prof. Timo Muotkaa mielenkiintoisesta aiheesta, asiantuntevasta ja laadukkaasta ohjauksesta sekä vaativista kommenteista työni kirjotusprosessin aikana, mitkä saivat minut työskentelemään kovemmin. Erityiskiitokset Jarnolle avustuksesta pohjaeläinten määrityksessä ja aineiston tilastollisessa analysoinnissa. Lisäksi haluan kiittää Tiina Nokelaa ohjauksesta ja avustuksesta molekyylibiologisten menetelmien kanssa sekä FT Mikko Tolkkista QIIMEanalyysien toteutuksesta. Kiitos myös Suomen ympäristökeskukselle työskentelytilojen mahdollistamisesta sekä taloudellisesta kannustuksesta. Lopuksi haluan kiittää perhettäni, ystäviäni ja opiskelutovereitani kaikesta läsnäolosta, tuesta ja kannustuksesta koko graduprojektini ajan. Erityiskiitokset Emmille, Pirjolle, Hannalle ja Niinalle vertaistuesta, sympatiasta sekä piristyksestä viimeisinä pitkinä kuukausina! 40
43 KIRJALLISUUS Ahtiainen, M. & P. Huttunen Metsätaloustoimenpiteiden pitkäaikaisvaikutukset purovesien laatuun ja kuormaan. Teoksessa: Saukkonen, S. & K. Kenttämies (toim.) Metsätalouden vesistövaikutukset ja niiden torjunta. METVE projektin loppuraportti. Suomen ympäristö 2: Ahvonen, A., E. Jutila, T. Järvenpää, A. Lappalainen, M. Rask & P. Vuorinen Metsätalouden vaikutukset kaloihin, rapuihin ja kalatalouteen. Kirjallisuusselvitys. Kalatutkimuksia s. RKTL. Helsinki. Allan, J.D Landscapes and riverscapes: The Influence of Land Use on Stream Ecosystems. Annual Review of Ecology, Evolution and Systematics 35: Allan, J.D. & M.M. Castillo Stream ecology: Structure and function of running waters. Springer, Netherlands. 436 s. Altschul, S.F., W. Gish, W. Miller, E. W. Myers & D. J. Lipman Basic local alignment search tool. Journal of Molecular Biology 215 (3): Anderson, M.J A New method for non parametric multivariate analysis of variance. Austral Ecology 26: Apitz S.E., M. Elliott, M. Fountain & T.S. Galloway European Environmental Management: Moving to an Ecosystem Approach. Integrated Environmental Assessment and Management 2: Bergfur, J., R.K. Johnson, L. Sandin, W. Goedkoop & K. Nygren Effects of nutrient enrichment on boreal streams: invertebrates, fungi and leaf litter breakdown. Freshwater Biology 52: Bergfur, J. & C. Sundberg Leaf litter associated fungi and bacteria along temporal and environmental gradients in boreal streams. Aquatic Microbial Ecology 73 (3): Boulton, A.J., & P.I. Boon A review of methodology used to measure leaf litter decomposition in lotic environments: time to turn over a new leaf? Australian Journal of Marine and Freshwater Research 42: Benfield, E.F Leaf breakdown in stream ecosystems. Teoksessa: Methods in Stream Ecology (Toim. R.F. Hauer & G.A. Lamberti), s Academic Press, San Diego, U.S.A. Benfield, E.F., J.R. Webster, J.L. Tank & J.J. Hutchens Long-Term Patterns in Leaf Breakdown in Streams in Response to Watershed Logging. International Review of Hydrobiology 86 (4 5): Bärlocher, F Molecular approaches promise a deeper and broader understanding of the evolutionary ecology of aquatic hyphomycetes. Journal of the North American Benthological Society 29:
44 Bärlocker, F Aquatic hyphomycetes in a changing environment. Fungal Ecology 19: Caporaso, J.G., J. Kuczynski, J. Stombaugh, K. Bittinger, F.D. Bushman, E.K. Costello, N. Fierer, A.G. Pena, J.K. Goodrich, J.I. Gordon, G.A. Huttley, S.T. Kelley, D. Knights, J.E. Koenig, R.E. Ley, C.A. Lozupone, D. McDonald, B.D. Muegge, M. Pirrung, J. Reeder, J.R. Sevinsky, P.J. Turnbaugh, W.A. Walters, J. Widmann, T. Yatsunenko, J. Zaneveld & R. Knight QIIME allows analysis of high throughput community sequencing data. Nature Methods 7 (5): Cummins, K.W Trophic relations of aquatic insects. Annual Review of Entomology 18: Cornut, J., A. Elger, D. Lambricot, P. Marmonier & E. Chauvet Early stages of leaf decomposition are mediated by aquatic fungi in the hyporheic zone of woodland streams. Freshwater Biology 55: Costantini, M.L. & L. Rossi Species diversity and decomposition in laboratory aquatic systems: the role of species interactions. Freshwater Biology 55: Danger, M., J. Cornut, A. Elger & E. Chauvet Effects of burial on leaf litter quality, microbial conditioning and palatability to three shredder taxa. Freshwater Biology 57: Dangles, O. & B. Malmqvist Species richness decomposition relationships depend on species dominance. Ecology Letters 7: Desjardins, P. & D. Conklin NanoDrop Microvolume Quantitation of Nucleic Acids. Journal of Visualized Experiments (45): e2565. Dudgeon, D., A.H. Arthington, M.O. Gessner, Z. I. Kawabata, D.J. Knowler, C. Lévêque, R.J. Naiman, A. H. Prieur Richard, D.Soto, M.L.J. Stiassny & C.A. Sullivan Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews 81: Dufrene, M. & P. Legendre Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67(3): Edgar, R.C Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26: Elosegi, A., J. Díez, & M. Murz Effects of hydromorphological integrity on biodiversity and functioning of river ecosystems. Hydrobiologia 657: van Emden, H. F Statistics for terrified biologists. Blacwell Publishing Ltd. 343 s. Euroopan parlamentti ja neuvosto Euroopan parlamentin ja neuvoston direktiivi 2000/60/EY, yhteisön vesienhoidon puitteista. Euroopan yhteisön virallinen lehti 327:
45 Feld, C.K, S. Birk, C.D. Bradley, D. Hering, J. Kail, A. Marzin, A. Mellcher, D. Nemitz, M.L. Pedersen, F. Pletterbauer,D. Point, P.F.M. Verdonschot & N. Frobergn From Natural to Degraded Rivers and Back Again: A Test of Restoration Ecology Theory and Practice. Advances in Ecological Research 44: Flores, L., A. Larranãga, J. Díez & A. Elosegi, Experimental wood addition in streams: effects on organic matter storage and breakdown. Freshwater Biology 56: Flushberg, B.A., D.R. Webster, J.H. Lee, K.J. Travers, E.C. Olivares, T.A. Clark, J. Korlach & S.W. Turner Direct detection of DNA methylation during single molecule, realtime sequencing. Nature Methods 7 (6): Gessner, M.O. & E. Chauvet A case for using litter breakdown to assess functional stream integrity. Ecological Applications 12 (2): Gessner, M. O., E. Chauvet & M. Dobson A Perspective on Leaf Litter Breakdown in Streams. Oikos 85 (2): Gessner, M.O., C.M. Swan, C.K. Dang, B.G. McKie, R.D. Bardgett, D.H. Wall & S. Hattenschwiler Diversity meets decomposition. Trends in Ecology & Evolution 25: Graça, M.A The Role of Invertebrates on Leaf Litter Decomposition in Streams a Review. International Review of Hydrobiology 86 (4 5): Haase, P., D. Hering, S.C. Jähnig, A.W. Lorenz & A. Sundermann The impact of hydromorphological restoration on river ecological status: a comparison of fish, benthic invertebrates, and macrophytes. Hydrobiologia 704: Heino, J., J. Ilmonen, J. Kotonen, H. Mykrä, L. Paasivirta, J. Soininen & R. Virtanen Surveying biodiversity in protected and managed areas: Algae, macrophytes and macroinvertebrates in boreal forest streams. Ecological Indicators 9 (6): Heino J. & A. Juntunen Kyröjoen vesistön koskien pohjaeläimistön vaihtelu ja veden laatu. Länsi-Suomen ympäristökeskus, moniste. 38 s. Heino, J., P. Louhi & T. Muotka Identifying the scales of variability in stream macroinvertebrate abundance, functional composition and assemblage structure. Freshwater Biology 49: Heino, J., T. Nokela, J. Soininen, M. Tolkkinen, L. Virtanen & R. Virtanen Elements of metacommunity structure and community environment relationships in stream organisms. Freshwater Biology 60: Heino J., M. Tolkkinen, A M. Pirttilä, H. Aisala & H. Mykrä Microbial diversity and community environment relationships in boreal streams. Journal of Biogeography 41:
46 Hering D., A. Borja, J. Carstensen, L. Carvalho, M. Elliot, C.K. Feld, A. Heiskanen, R.K. Johnson, J. Moe, D. Pont, A.L. Solheim & W. van de Bund The European Water Framework Directive at the age of 10: A critical review of the achievements with recommendations for the future. The Science of the Total Environment 408: Hieber, M. & M.O. Gessner Contribution of stream detrivores, fungi, and bacteria to leaf breakdown based on biomass estimates. Ecology 83 (4): Hill, W.R Factors affecting benthic algae effects of light. Teoksessa: Algal Ecology Freshwater Benthic Ecosystems. (Toim. R.J. Stevenson, M.L. Bothwell & R.L. Lowe). Academic Press, San Diego, U.S.A. 788 s. Holopainen, M. & P. Pulkkinen Tilastolliset menetelmät. Werner Söderström Osakeyhtiö, Helsinki. 338 s. Huttunen, K. L., H. Mykrä & T. Muotka Temporal variation in taxonomic completeness of stream macroinvertebrate assemblages. Freshwater Science 31: Jones, J.I., J.F. Murphy, A.L. Collins, D.A. Sear, P.S. Naden & P.D. Armitage The impact of fine sediment on macroinvertebrates. River Research and Applications 28: Jonsson, M., B. Malmqvist & P. O. Hoffsten Leaf litter breakdown rates in boreal streams: does shredder species richness matter? Freshwater Biology 46: Jähning, S.C., A.W. Lorenz & D. Hering Re meandering German lowland streams: Qualitative and quantitative effects of restoration measures on hydromorphology and macroinvertebrates. Environmental Management 44: Kemp, P., D. Sear, A. Collins, P. Naden & I. Jones The impacts of fine sediment on riverine fish. Hydrological Processes 25: Kenttämies. K. & T. Mattsson (toim.) Metsätalouden vesistökuormitus MESUVE-projektin loppuraportti. Suomen ympäristö 816. Suomen ympäristökeskus, Helsinki. 160 s. Kuehn, K.A Lentic and lotic habitats as templets for fungal communities: traits, adaptations, and their significance to litter decomposition within freshwater ecosystems. Fungal Ecology 19: Lake, P. S Disturbance, patchiness and diversity in streams. Journal of American Benthological Society 19 (4): Lake, P.S., N. Bond & P. Reich Linking ecological theory with stream restoration. Freshwater Biology 52: Lecerf, A. & E. Chauvet Diversity and functions of leaf decaying fungi in human altered stream. Freshwater Biology 53 (8):
47 Lepori, F., D. Palm & B. Malmqvist. 2005a. Effects of stream restoration on ecosystem functioning: detritus retentiveness and decomposition. Journal of Applied Ecology 42: Lepori, F., D. Palm, E. Brännäs & B. Malmqvist. 2005b. Does restoration of structural heterogeneity in streams enchange fish and macroinvertebrate diversity? Ecological applications 15(6): Leps, M., A Sundermann, J. D. Tonkin, A.W. Lorenz & P. Haase Time is no healer: increasing restoration age does not lead to improved benthic invertebrate communities in restored river reaches. Science of the Total Environment : Lillehammer, A Norwegian stoneflies. IV. Laboratory studies on ecological factors influencing distribution. Norwegian. Journal of Entomology 22 (2): Lorenz, A.W. & C.K. Feld Upstream river morphology and riparian land use overrule local restoration effects on ecological status assessment. Hydrobiologia 704: Louhi, P., H. Mykrä, R. Paavola, A. Huusko, T. Vehanen, A. Mäki Petäys & T. Muotka Twenty years of stream restoration in Finland: Little response by benthic macroinvertebrate communities. A Review. Ecological Applications 21 (6): Madigan, M.T., J.M. Martunko, P.V. Dunlap & D.P. Clark Biology of microorganisms. 12. painos. Pearson Benjamin Cummings, San Fransisco, USA s. Martel, M., M.A. Rodríguez, & P. Bérubé Multi scale analysis of responses of stream macrobenthos to forestry activities and environmental context. Freshwater Biology 52: Marttila, M., P. Louhi, A. Huusko, A. Mäki Petäys, T. Yrjänä & T. Muotka Longterm performance of in-stream restoration measures in boreal streams. Ecohydrology 9 (2): Matthaei, C.D., J.J. Piggott & C.R. Townsend Multiple stressors in agricultural streams: interactions among sediment addition, nutrient enrichment and water abstraction. Journal of Applied Ecology 47(3): McCune B. & J. B. Grace Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon, 300 s. McKie, B.G. & B. Malqvist Assessing ecosystem functioning in streams affected by forest management: increased leaf decomposition occurs without changes to the composition of benthic assemblages. Freshwater Biology 54: Miller, S.W., P. Budy & J.C. Schmidt Quantifying macroinvertebrate responses to in-stream habitat restoration: applications of meta analysis to river restoration. Restoration Ecology 18:
48 Moog O. (Toim.) Fauna Aquatica Austriaca, Edition Wasserwirtschaftsskataster, Bundesministerium für Land und Forstwirtschaft, Umwelt und Wasserwirtschaft, Vienna. Muotka, T. & P. Laasonen Ecosystem recovery in restored headwater streams: The role of enhanced leaf retention. Journal of Applied Ecology 39: Muotka, T., R. Paavola, A. Haapala, M. Novikmec & P. Laasonen Long term recovery of stream habitat structure and benthic invertebrate communities from in stream restoration. Biological Conservation 105: Muotka T. & J. Syrjänen Changes in habitat structure, benthic invertebrate diversity, trout populations and ecosystem processes in restored forest streams: a boreal perspective. Freshwater Biology 52: Newbury, R.W Hydrological determinants of aquatic insect habitats. Teoksessa: The Ecology of Aquatic Insects. Toim. V. H. Resh & D. M. Rosenberg. s Praeger, New York, USA. Owens, P.N., R.J. Batalla, A.J. Collins, B. Gomez, D.M. Hicks, A.J. Horowitz, G.M. Kondolf, M. Marden, M.J. Page, D.H. Peacock, E.L. Petticrew, W. Salomons & N.A. Trustrum Fine grained sediment in river systems: environmental significance and management issues. River Research and Applications 21: Palmer, M.A., R.F. Ambrose & N.L. Poff Ecological theory and community restoration ecology. Restoration Ecology 5 (4): Palmer, M.A., H.L. Mellinger, & E. Bernhardt River restoration, habitat heterogeneity and biodiversity: a failure of theory or practice? Freshwater Biology 55 (1): Peltomaa, R Drainage of forests in Finland. Irrigation & Drainage 56: Pfankuch, D.J Stream reach inventory and channel stability evaluation. U.S.D.A. Forest Service, Region 1, Missoula, Montana, U.S.A. Pickett, S.T.A. & P.S. White The ecology of natural disturbance and patch dynamics. Academic Press Inc., San Diego, California. 472 s. Poff, N.L., J.D. Allan, M.B. Bain, J.R. Karr, K.L. Prestegaard, B.D. Richter, R.E. Sparks & J.C. Stromberg The Natural Flow Regime: A paradigm for river conservation and restoration. BioScience 47 (11): Poff, N.L Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. Journal of American Benthological Society 16 (2): Raunio A., A. Schulman & T. Kontula (toim.) Suomen luontotyyppien uhanalaisuus Osa 1: Tulokset ja arvioinnin perusteet. Suomen ympäristö 8/2008. Suomen ympäristökeskus, Helsinki. 264 s. 46
49 Resh, V.H., A.V. Brown, A.P. Covich, M.E. Gurtz, H.W. Li, G.W. Minshall, S.R. Reice, A.L. Sheldon, J.B. Wallace & R.C. Wissmar The role of disturbance in stream ecology. Journal of American Benthological Society 7 (4): Richards, C., R.J. Haro, L.B. Johnson & G.E. Host Catchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshwater Biology 37 (1): Roni, P., T. Beechie, G. Press & K. Hanson Wood placement in river restoration: fact, fiction, and future direction. Canadian Journal of Fisheries and Aquatic Sciences 72: Roni, P., K. Hanson & T. Beechie Global review of the physical and biological effectiveness of stream habitat rehabilitation techniques. North American Journal of Fisheries Management 28: Sanpera-Calbet, I., E. Chauvet & J.S. Richardson Fine sediment on leaves: shredder removal of sediment does not enhance fungal colonisation. Aquatic Sciences 74: Sundermann, A., S. Stoll & P. Haase River restoration success depends on the species pool of the immediate surroundings. Ecological Applications 21: Sutela, T., M. Olin, T. Vehanen & M. Rask Hajakuormituksen vaikutukset järvien ja jokien kalastoon ja ekologiseen tilaan. Kala- ja riistaraportteja 411. Riista- ja kalatalouden tutkimuslaitos, Helsinki, 36 s. Sutherland, A.B., J.M. Culp & G.A. Benoy Evaluation of deposited sediment and macroinvertebrate metrics used to quantify biological response to excessive sedimentation in agricultural streams. Environmental Management 50: Stewart Oaten, A., W.M. Murdoch & K.R. Parker, Environmental impact assessment:"pseudoreplication" in time? Ecology 67: Tolkkinen M., H. Mykrä, AM. Markkola, H. Aisala, K M. Vuori, J. Lumme, A M. Pirttilä & T. Muotka Decomposer communities in human-impacted streams: species dominance rather than richness affects leaf decomposition. Journal of Applied Ecology Tolkkinen M.J., H. Mykrä, R. Virtanen, M. Tolkkinen, T. Kauppila, L. Paasivirta & T. Muotka Land use impacts on stream community composition and concordance along a natural stress gradient. Ecological Indicators 62: Underwood, A On beyond BACI: sampling designs that might reliably detect environmental disturbances. Ecological Applications 4: Vannote R.L., G.W. Minshall, K.W. Cummins, J.R. Sedell & C.E. Cushing The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37:
50 Vaughan, I.P., M. Diamond, A.M. Gurnell, K.A. Hall, A. Jenkins, N.J. Milner, L.A. Naylor, D.A. Sear, G. Woodward & S.J. Ormerod Integrating ecology with hydromorphology: A priority for river science and management. Aquatic Conservation: Marine and Freshwater Ecosystems 19: Wallace, J.B., J.W. Grubaugh & M.R. Whiles Influence of Coarse Woody Debris on Stream Habitats and Invertebrate Biodiversity. U.S. Department of Agriculture, Forest Service, Southern Research Station. General Technical Report SE 94: Wallace, J.B. & J.R. Webster The role of macroinvertebrates in stream ecosystem function. Annual Review of Entomology 19 (41): Wallace, J.B., S.L. Eggert, J.L. Meyer & J.R. Webster Stream invertebrate productivity linked to forest subsidies: 37 stream-years of reference and experimental data. Ecology 96 (5): Webster, J.R, E.F. Benfield, T.P. Ehrman, M.A. Schaeffer, J.L. Tank, J.J. Huthens & D.J. D Angelo What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. Freshwater Biology 41: Webster, J.R. & E.F. Benfield Vascular plat breakdown in freshwater ecosystems. Annual Review of Ecology, Evolution, and Systematics 17: Wood, P.J. & P.D. Armitage Biological Effects of Fine Sediment in the Lotic Environment. Environmental Management 21 (2): Vuori, K.-M. & I. Joensuu Impact of forest drainage on the macroinvertebrates of small boreal headwater stream: Do buffer zones protect lotic biodiversity? Biological Conservation 77: Vuori, K.-M., I. Joensuu, J. Latvala, E. Jutila & A. Ahvonen Forest drainage: a threat to benthic biodiversity of boreal headwater streams? Aquatic Conservation: Marine and Freshwater Ecosystems 8: Wentworth, C.K A scale of grade and class terms for clastic sediments. The Journal of Geology 30 (5): Yoshimura, M Effects of forest disturbances on aquatic insect assemblages. Entomological Science 15: Young, R.G., C.D. Matthaei & C.R. Townsend Organic matter breakdown and ecosystem metabolism: functional indicators for assessing river ecosystem health. Journal of American Benthological Society 27(3): Yrjänä, T., P.-L. Luhta, E. Hartikainen, E. Moilanen, S. Tammela, H. Marttila, B. Klöve, H. Suurkuukka, R. Virtanen & T. Muotka Liettyneiden metsäpurojen kunnostaminen. Metsätieteen aikakauskirja 2:
51 Zegling, L.H Stream microbial diversity in response to environmental changes: review and synthesis of existing research. Frontiers in Microbiology 6: 454. Internet-lähteet L /1299. Laki vesienhoidon ja merenhoidon järjestämisestä. Viitattu Saatavilla: < Life Technologies Corporation Ion Torrent Semiconductor sequencing technology overview. Viitattu Saatavilla: < Oksanen, J., F.G. Blanchet, R. Kindt, P. Legendre, P.R. Minchin, R.B. O'Hara, G. Simpson, P. Solymos, M.H.H. Stevens & H. Wagner. (2016). Vegan: Community Ecology Package. R package version Viitattu Saatavilla: < Meissner, K., J. Aroviita, S. Hellsten, M. Järvinen, S-M. Karjalainen, M. Kuoppala, H. Mykrä ja K-M. Vuori. Jokien ja järvien biologinen seuranta näytteenotosta tiedon tallentamiseen. Saatavilla: < gisten_seurantamenetelmien_ohjeet> MO BIO Laboratories PowerSoil DNA Isolation Kit, Instruction Manual. Version: Viitattu Saatavilla: < R Core Team The R Stats Package. R Package version Viitattu Saatavilla: < Ripley B.D. Choose a model by AIC in a Stepwise Algorithm. R Documentation. Saatavilla: < Roberts D.W Labdsv: Ordination and Multivariate Analysis for Ecology. R package version Viitattu Saatavilla: < 49

52 Liite 1. Tutkimuspurojen ympäristömuuttuja- ja maankäyttöaineisto. REF = Luonnontilainen puro, IMP = hiekoittunut puro, REST = kunnostettu puro.
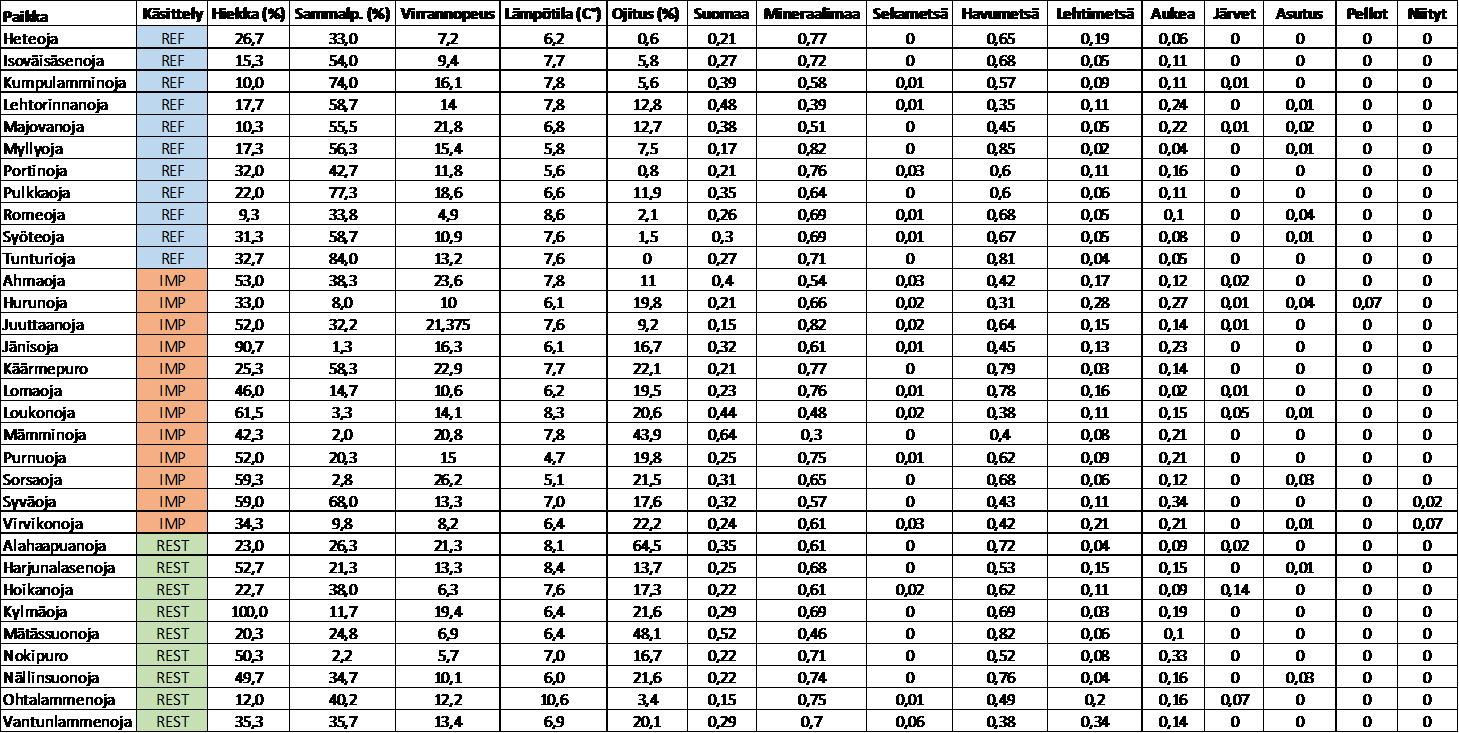
53 51

54 Liite 2. Tutkimuspaikoilla tavatut pohjaeläintaksonit ja niiden ravinnonkäyttöryhmät (FFG). PRED = peto, GRAZ = laiduntaja, SH = pilkkoja, FILT = suodattaja. Sinisellä kuvattu luonnontilaiset, oranssilla hiekoittuneet ja vihreällä kunnostetut purot. 52
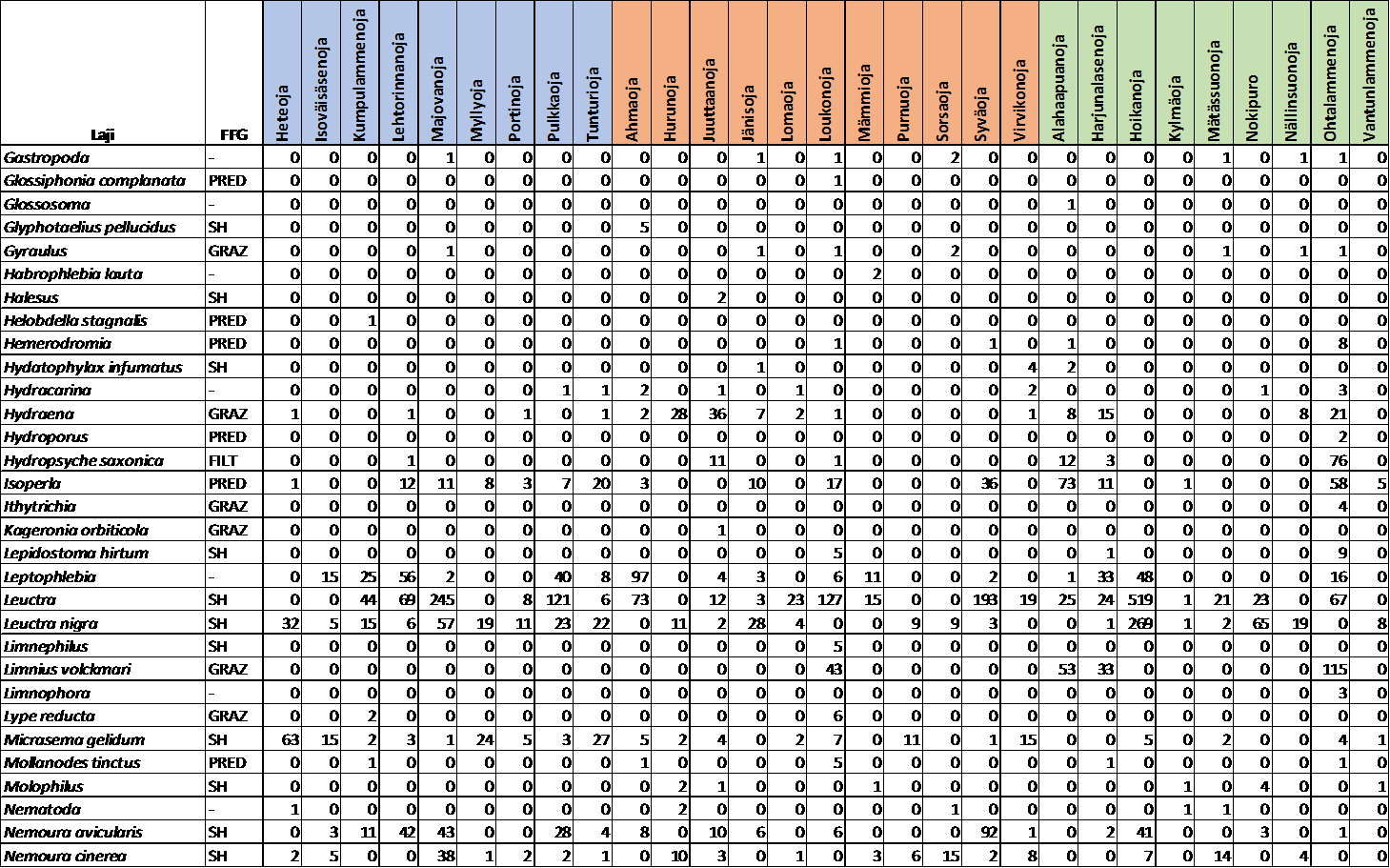
55 53
Metsäpurojen kunnostamisen hydrauliset vaikutukset
 Metsäpurojen kunnostamisen hydrauliset vaikutukset Vesistökunnostusverkoston vuosiseminaari 2016 Hannu Marttila Vesi- ja ympäristötekniikan tutkimusryhmä Oulun yliopisto Latvavesiä on muokattu Suomessa
Metsäpurojen kunnostamisen hydrauliset vaikutukset Vesistökunnostusverkoston vuosiseminaari 2016 Hannu Marttila Vesi- ja ympäristötekniikan tutkimusryhmä Oulun yliopisto Latvavesiä on muokattu Suomessa
Metsäpurojen kunnostusten vaikutukset ekosysteemien rakenteeseen ja toimintaan: ReFFECT-hankkeen tuloksia
 Metsäpurojen kunnostusten vaikutukset ekosysteemien rakenteeseen ja toimintaan: ReFFECT-hankkeen tuloksia, SYKE, Vesikeskus OY: Timo Muotka, Pauliina Louhi, Björn Klöve, Hannu Marttila, Simo Tammela SYKE:,
Metsäpurojen kunnostusten vaikutukset ekosysteemien rakenteeseen ja toimintaan: ReFFECT-hankkeen tuloksia, SYKE, Vesikeskus OY: Timo Muotka, Pauliina Louhi, Björn Klöve, Hannu Marttila, Simo Tammela SYKE:,
Kiintoainemenetelmien käyttö turvemaiden alapuolella. Hannu Marttila
 Kiintoainemenetelmien käyttö turvemaiden alapuolella Hannu Marttila Motivaatio Orgaaninen kiintoaines ja sedimentti Lisääntynyt kulkeutuminen johtuen maankäytöstä. Ongelmallinen etenkin turvemailla, missä
Kiintoainemenetelmien käyttö turvemaiden alapuolella Hannu Marttila Motivaatio Orgaaninen kiintoaines ja sedimentti Lisääntynyt kulkeutuminen johtuen maankäytöstä. Ongelmallinen etenkin turvemailla, missä
Joen määritelmä. Joella tarkoitetaan virtaavan veden vesistöä. Joen valuma-alue on vähintään 100 km 2.
 Joet ja kunnostus Joen määritelmä Joella tarkoitetaan virtaavan veden vesistöä. Joen valuma-alue on vähintään 100 km 2. Valuma-alueella tarkoitetaan aluetta, jolta vedet kerääntyvät samaan vesistöön. Jokiekosysteemin
Joet ja kunnostus Joen määritelmä Joella tarkoitetaan virtaavan veden vesistöä. Joen valuma-alue on vähintään 100 km 2. Valuma-alueella tarkoitetaan aluetta, jolta vedet kerääntyvät samaan vesistöön. Jokiekosysteemin
Metsäpurojen rantavyöhykkeet monimuotoisuuden lähteinä. Jarno Turunen & Mari Tolkkinen Suomen ympäristökeskus (SYKE)
") Metsäpurojen rantavyöhykkeet monimuotoisuuden lähteinä Jarno Turunen & Mari Tolkkinen Suomen ympäristökeskus (SYKE) 13.6.2018 Metsäpuron rantavyöhyke = puron vaikutuspiirissä oleva metsäalue Metsä- ja
Metsäpurojen rantavyöhykkeet monimuotoisuuden lähteinä Jarno Turunen & Mari Tolkkinen Suomen ympäristökeskus (SYKE) 13.6.2018 Metsäpuron rantavyöhyke = puron vaikutuspiirissä oleva metsäalue Metsä- ja
Opetusmateriaali on tuotettu osana vesistökunnostusverkoston toimintaa ja on vapaasti kaikkien käytettävissä ja muokattavissa.
 Opetusmateriaali on tuotettu osana vesistökunnostusverkoston toimintaa ja on vapaasti kaikkien käytettävissä ja muokattavissa. Jokien kunnostus Sisällysluettelo Tiesitkö tämän joesta - Jokien tietopaketti
Opetusmateriaali on tuotettu osana vesistökunnostusverkoston toimintaa ja on vapaasti kaikkien käytettävissä ja muokattavissa. Jokien kunnostus Sisällysluettelo Tiesitkö tämän joesta - Jokien tietopaketti
Tiina Tulonen Lammin biologinen asema Helsingin yliopisto
 Tiina Tulonen Lammin biologinen asema Helsingin yliopisto Kokonaiskuormituksesta hajakuormituksen osuus on fosforin osalta n. 60 % ja typen osalta n 80% (SYKE tilastot) Fosfori Typpi Toimenpiteiden kohdentaminen
Tiina Tulonen Lammin biologinen asema Helsingin yliopisto Kokonaiskuormituksesta hajakuormituksen osuus on fosforin osalta n. 60 % ja typen osalta n 80% (SYKE tilastot) Fosfori Typpi Toimenpiteiden kohdentaminen
Heinijärven vedenlaatuselvitys 2014
 Heinijärven vedenlaatuselvitys 2014 Tiina Tulonen Lammin biologinen asema Helsingin yliopisto 3.12.2014 Johdanto Heinijärven ja siihen laskevien ojien vedenlaatua selvitettiin vuonna 2014 Helsingin yliopiston
Heinijärven vedenlaatuselvitys 2014 Tiina Tulonen Lammin biologinen asema Helsingin yliopisto 3.12.2014 Johdanto Heinijärven ja siihen laskevien ojien vedenlaatua selvitettiin vuonna 2014 Helsingin yliopiston
TUULOKSEN PANNUJÄRVEN TILAN KEHITYS SEDIMENTIN PIILEVÄANA-
 TUULOKSEN PANNUJÄRVEN TILAN KEHITYS SEDIMENTIN PIILEVÄANA- LYYSIEN VALOSSA Järvi Hämeenlinnan Tuuloksen Pannujärvi (tunnus 35.793.1.002, vesiala 0,362 km 2, suurin syvyys 12 m ja tilavuus 1,4 milj. m 3
TUULOKSEN PANNUJÄRVEN TILAN KEHITYS SEDIMENTIN PIILEVÄANA- LYYSIEN VALOSSA Järvi Hämeenlinnan Tuuloksen Pannujärvi (tunnus 35.793.1.002, vesiala 0,362 km 2, suurin syvyys 12 m ja tilavuus 1,4 milj. m 3
PCR - tekniikka elintarvikeanalytiikassa
 PCR - tekniikka elintarvikeanalytiikassa Listerian, Salmonellan ja kampylobakteerien tunnistus elintarvikkeista ja rehuista 29.11.2012 Eva Fredriksson-Lidsle Listeria monocytogenes Salmonella (spp) Campylobacter
PCR - tekniikka elintarvikeanalytiikassa Listerian, Salmonellan ja kampylobakteerien tunnistus elintarvikkeista ja rehuista 29.11.2012 Eva Fredriksson-Lidsle Listeria monocytogenes Salmonella (spp) Campylobacter
Metsätalouden vaikutukset Kitkaja Posionjärvien tilaan
 Metsätalouden vaikutukset Kitkaja Posionjärvien tilaan Keskustelutilaisuus metsänomistajille 16.12.2014 Nuorisokeskus Oivanki Kati Häkkilä & Teemu Ulvi, SYKE Järvien tilassa havaittu muutoksia Asukkaat
Metsätalouden vaikutukset Kitkaja Posionjärvien tilaan Keskustelutilaisuus metsänomistajille 16.12.2014 Nuorisokeskus Oivanki Kati Häkkilä & Teemu Ulvi, SYKE Järvien tilassa havaittu muutoksia Asukkaat
Kosteikot leikkaavat ravinnekuormitusta ja elävöittävät maisemaa
 Liite 17.12.2007 64. vuosikerta Numero 3 Sivu 5 Kosteikot leikkaavat ravinnekuormitusta ja elävöittävät maisemaa Markku Puustinen, Suomen ympäristökeskus Kosteikot pidättävät tehokkaasti pelloilta valtaojiin
Liite 17.12.2007 64. vuosikerta Numero 3 Sivu 5 Kosteikot leikkaavat ravinnekuormitusta ja elävöittävät maisemaa Markku Puustinen, Suomen ympäristökeskus Kosteikot pidättävät tehokkaasti pelloilta valtaojiin
Paikkatiedon hyödyntämismahdollisuudet pienvesien tilan ja kunnostustarpeen arvioinnissa
 Paikkatiedon hyödyntämismahdollisuudet pienvesien tilan ja kunnostustarpeen arvioinnissa Teemu Ulvi, SYKE Metsätalouden vesiensuojelupäivät, Koli 22.9.2015 Sisältö Purojen tila arviointimenetelmien tarve
Paikkatiedon hyödyntämismahdollisuudet pienvesien tilan ja kunnostustarpeen arvioinnissa Teemu Ulvi, SYKE Metsätalouden vesiensuojelupäivät, Koli 22.9.2015 Sisältö Purojen tila arviointimenetelmien tarve
Metsätalouden ekologiset vesistövaikutukset: Pienvesien tilan seuranta ja luokittelun kehitystarpeet
 Metsätalouden ekologiset vesistövaikutukset: Pienvesien tilan seuranta ja luokittelun kehitystarpeet Jaana Rääpysjärvi, Jukka Aroviita, Satu Maaria Karjalainen, Minna Kuoppala, Jarno Turunen Suomen ympäristökeskus
Metsätalouden ekologiset vesistövaikutukset: Pienvesien tilan seuranta ja luokittelun kehitystarpeet Jaana Rääpysjärvi, Jukka Aroviita, Satu Maaria Karjalainen, Minna Kuoppala, Jarno Turunen Suomen ympäristökeskus
S A V O K A R J A L A N Y M P Ä R I S T Ö T U T K I M U S O Y
 S A V O K A R J A L A N Y M P Ä R I S T Ö T U T K I M U S O Y Linnunmaa Oy/ Tiia Grönholm (email) E 5141 28.12.2011 Lähetämme ohessa Endomines Oy:n uusien hankealueiden taustatilanteen selvittämiseen kuuluvan
S A V O K A R J A L A N Y M P Ä R I S T Ö T U T K I M U S O Y Linnunmaa Oy/ Tiia Grönholm (email) E 5141 28.12.2011 Lähetämme ohessa Endomines Oy:n uusien hankealueiden taustatilanteen selvittämiseen kuuluvan
Tornionjoen Suomen puoleisten pintavesien luokittelu ja ehdotetut lisätoimenpiteet
 Tornionjoen Suomen puoleisten pintavesien luokittelu ja ehdotetut lisätoimenpiteet Petri Liljaniemi Biologi Lapin ympäristökeskus 1 Vesistön ekologisen tilan luokittelu Biologiset tekijät Levät, vesikasvillisuus,
Tornionjoen Suomen puoleisten pintavesien luokittelu ja ehdotetut lisätoimenpiteet Petri Liljaniemi Biologi Lapin ympäristökeskus 1 Vesistön ekologisen tilan luokittelu Biologiset tekijät Levät, vesikasvillisuus,
Virtavesikunnostuksien ekologinen vaikuttavuus
 Virtavesikunnostuksien ekologinen vaikuttavuus Kalatalouden ympäristöohjelma Pauliina Louhi Erikoistutkija pauliina.louhi@luke.fi Koskikunnostuksia jo 1970-luvun lopulta lähtien Tukinuitto ja muut perkaukset,
Virtavesikunnostuksien ekologinen vaikuttavuus Kalatalouden ympäristöohjelma Pauliina Louhi Erikoistutkija pauliina.louhi@luke.fi Koskikunnostuksia jo 1970-luvun lopulta lähtien Tukinuitto ja muut perkaukset,
TAIMENEN KUTUPESÄINVENTOINTI
 Vesi-Visio Visio osk Opettajantie 7-9 B15 40900 SÄYNÄTSALO www.vesi-visio.netvisio.net +35840-7030098 TAIMENEN KUTUPESÄINVENTOINTI Mitä, miksi, miten, milloin? Tietoa ja ohjeistusta toiminnasta ja käytännön
Vesi-Visio Visio osk Opettajantie 7-9 B15 40900 SÄYNÄTSALO www.vesi-visio.netvisio.net +35840-7030098 TAIMENEN KUTUPESÄINVENTOINTI Mitä, miksi, miten, milloin? Tietoa ja ohjeistusta toiminnasta ja käytännön
Keliberin kaivoshankkeen perustilaselvitys
 Keliberin kaivoshankkeen perustilaselvitys Piileväselvitys kesällä 2014 Selvityksessä tutkittiin suunnittelualueen vesien nykytila piileväanalyysien avulla viidellä havaintopaikalla. Piileväanalyysit Näytteenotossa,
Keliberin kaivoshankkeen perustilaselvitys Piileväselvitys kesällä 2014 Selvityksessä tutkittiin suunnittelualueen vesien nykytila piileväanalyysien avulla viidellä havaintopaikalla. Piileväanalyysit Näytteenotossa,
Itämeren sedimentin ja rautamangaanisaostumien. hajottaa raakaöljyä ja naftaleenia. Suomen ympäristökeskus
 Itämeren sedimentin ja rautamangaanisaostumien bakteerien kyky hajottaa raakaöljyä ja naftaleenia Mikrokosmoskokeet 23.7.-18.12.2012 Anna Reunamo, Pirjo Yli-Hemminki, Jari Nuutinen, Jouni Lehtoranta, Kirsten
Itämeren sedimentin ja rautamangaanisaostumien bakteerien kyky hajottaa raakaöljyä ja naftaleenia Mikrokosmoskokeet 23.7.-18.12.2012 Anna Reunamo, Pirjo Yli-Hemminki, Jari Nuutinen, Jouni Lehtoranta, Kirsten
Vesiekosysteemien kestävä kunnostus. ReEFFECT ja AQUADIGM
 Vesiekosysteemien kestävä kunnostus ReEFFECT ja AQUADIGM Vesiekosysteemien kestävä hallinta edellyttää ihmistoiminnan vaikutusten kontrollointia Hoito Kunnostus ReEFFECT Virtavedet Björn Klöve, Oulun yliopisto
Vesiekosysteemien kestävä kunnostus ReEFFECT ja AQUADIGM Vesiekosysteemien kestävä hallinta edellyttää ihmistoiminnan vaikutusten kontrollointia Hoito Kunnostus ReEFFECT Virtavedet Björn Klöve, Oulun yliopisto
MIKSI JÄRVI SAIRASTUU?
 Pekka Sojakka Etelä-Savon ELY-keskus MIKSI JÄRVI SAIRASTUU? MIKÄ ON KOTIJÄRVENI TILA? PÄÄTEEMA: REHEVÖITYMINEN KÄSITTEET REHEVÖITYMINEN(eutrofoituminen) -REHEVÖITYMINEN on yksi LIKAANTUMISEN ja PILAANTUMISEN
Pekka Sojakka Etelä-Savon ELY-keskus MIKSI JÄRVI SAIRASTUU? MIKÄ ON KOTIJÄRVENI TILA? PÄÄTEEMA: REHEVÖITYMINEN KÄSITTEET REHEVÖITYMINEN(eutrofoituminen) -REHEVÖITYMINEN on yksi LIKAANTUMISEN ja PILAANTUMISEN
ISO-KAIRIN VEDEN LAATU Kesän 2015 tutkimus ja vertailu vuosiin 1978, 1980 ja 1992
 LUVY/149 4.8.215 Minna Sulander Ympäristönsuojelu, Vihti ISO-KAIRIN VEDEN LAATU Kesän 215 tutkimus ja vertailu vuosiin 1978, 198 ja 1992 Vihdin pohjoisosassa sijaitsevasta Iso-Kairista otettiin vesinäytteet
LUVY/149 4.8.215 Minna Sulander Ympäristönsuojelu, Vihti ISO-KAIRIN VEDEN LAATU Kesän 215 tutkimus ja vertailu vuosiin 1978, 198 ja 1992 Vihdin pohjoisosassa sijaitsevasta Iso-Kairista otettiin vesinäytteet
Surviaissääskien kotelonahkamenetelmä (CPET) Tulokset ja johtopäätökset
 Tulokset ja johtopäätökset") Surviaissääskien kotelonahkamenetelmä (CPET) Tulokset ja johtopäätökset Janne Raunio, FT, Kymijoen vesi ja ympäristö ry. Mika Nieminen, FM, SYKE / Jyväskylän yliopisto BioTar-loppuseminaari 14.5.2014,
Surviaissääskien kotelonahkamenetelmä (CPET) Tulokset ja johtopäätökset Janne Raunio, FT, Kymijoen vesi ja ympäristö ry. Mika Nieminen, FM, SYKE / Jyväskylän yliopisto BioTar-loppuseminaari 14.5.2014,
Tervetuloa retkelle! Kunnostettujen purojen ja rumpujen valtakuntaan Iijoen vesistöalueelle
 Tervetuloa retkelle! Kunnostettujen purojen ja rumpujen valtakuntaan Iijoen vesistöalueelle Pirkko-Liisa Luhta p.+358400 293023 pirkko-liisa.luhta@metsa.fi Eero Moilanen p.+35840 5314969 eero.t.moilanen@metsa.fi
Tervetuloa retkelle! Kunnostettujen purojen ja rumpujen valtakuntaan Iijoen vesistöalueelle Pirkko-Liisa Luhta p.+358400 293023 pirkko-liisa.luhta@metsa.fi Eero Moilanen p.+35840 5314969 eero.t.moilanen@metsa.fi
UUDET TEKNIIKAT SISÄYMPÄRISTÖN MIKROBIEN TOTEAMISESSA
 UUDET TEKNIIKAT SISÄYMPÄRISTÖN MIKROBIEN TOTEAMISESSA LIITU-päivä 4.5.2006 FT Helena Rintala Kansanterveyslaitos, Ympäristöterveyden osasto Mihin sisäympäristön mikrobien mittauksia tarvitaan? Rakennusten
UUDET TEKNIIKAT SISÄYMPÄRISTÖN MIKROBIEN TOTEAMISESSA LIITU-päivä 4.5.2006 FT Helena Rintala Kansanterveyslaitos, Ympäristöterveyden osasto Mihin sisäympäristön mikrobien mittauksia tarvitaan? Rakennusten
Vesiensuojelu metsätaloudessa Biotalous tänään ja huomenna Saarijärvi 28.1.2016. Juha Jämsén Suomen metsäkeskus
 Vesiensuojelu metsätaloudessa Biotalous tänään ja huomenna Saarijärvi 28.1.2016 Juha Jämsén Suomen metsäkeskus Metsätalouden vesistökuormitus Metsätalouden kuormitus on tyypiltään hajakuormitusta. Myös
Vesiensuojelu metsätaloudessa Biotalous tänään ja huomenna Saarijärvi 28.1.2016 Juha Jämsén Suomen metsäkeskus Metsätalouden vesistökuormitus Metsätalouden kuormitus on tyypiltään hajakuormitusta. Myös
Kolmen helmen joet hanke
 Hämeenkyrön kunta, Nokian kaupunki, Ylöjärven kaupunki Kolmen helmen joet hanke Virtavesi-inventointi ja kunnostussuunnitelma Rapujen istutuksen riskianalyysi 3.2.2017 Page 1 Rapujen istutuksen riskianalyysi
Hämeenkyrön kunta, Nokian kaupunki, Ylöjärven kaupunki Kolmen helmen joet hanke Virtavesi-inventointi ja kunnostussuunnitelma Rapujen istutuksen riskianalyysi 3.2.2017 Page 1 Rapujen istutuksen riskianalyysi
Asuinalueen rakentamisen vaikutukset veden laatuun, virtaamaan ja ainekuormitukseen - Esimerkkinä Espoon Suurpelto 2006-2012
 Asuinalueen rakentamisen vaikutukset veden laatuun, virtaamaan ja ainekuormitukseen - Esimerkkinä Espoon Suurpelto 2006-2012 Pienvesitapaaminen 2.6.2014 Päivi Haatainen Helsingin yliopisto Geotieteiden
Asuinalueen rakentamisen vaikutukset veden laatuun, virtaamaan ja ainekuormitukseen - Esimerkkinä Espoon Suurpelto 2006-2012 Pienvesitapaaminen 2.6.2014 Päivi Haatainen Helsingin yliopisto Geotieteiden
Kunnostusojituksen aiheuttama humuskuormitus Marjo Palviainen
 Kunnostusojituksen aiheuttama humuskuormitus Marjo Palviainen Maatalous-metsätieteellinen tiedekunta /Metsätieteiden laitos 10.10.2013 1 Kunnostusojitukset ja humuskuormitus Suomen soista yli puolet (54
Kunnostusojituksen aiheuttama humuskuormitus Marjo Palviainen Maatalous-metsätieteellinen tiedekunta /Metsätieteiden laitos 10.10.2013 1 Kunnostusojitukset ja humuskuormitus Suomen soista yli puolet (54
Hapetuksen tarkoitus purkamaan pohjalle kertyneitä orgaanisen aineksen ylijäämiä
 Hapetuksen tarkoitus Hapettamiselle voidaan asettaa joko lyhytaikainen tai pitkäaikainen tavoite: joko annetaan kaloille talvisin mahdollisuus selviytyä pahimman yli tai sitten pyritään hillitsemään järven
Hapetuksen tarkoitus Hapettamiselle voidaan asettaa joko lyhytaikainen tai pitkäaikainen tavoite: joko annetaan kaloille talvisin mahdollisuus selviytyä pahimman yli tai sitten pyritään hillitsemään järven
HANNUKAISEN KAIVOSHANKE TAIMENEN POTENTIAALISTEN LISÄÄNTYMISALUEIDEN SEL- VITYS
 Vastaanottaja Hannukainen Mining Oy Asiakirjatyyppi Muistio Päivämäärä 30.9.2016 HANNUKAISEN KAIVOSHANKE TAIMENEN POTENTIAALISTEN LISÄÄNTYMISALUEIDEN SEL- VITYS Päivämäärä 30.9.2016 Laatija Tarkastaja
Vastaanottaja Hannukainen Mining Oy Asiakirjatyyppi Muistio Päivämäärä 30.9.2016 HANNUKAISEN KAIVOSHANKE TAIMENEN POTENTIAALISTEN LISÄÄNTYMISALUEIDEN SEL- VITYS Päivämäärä 30.9.2016 Laatija Tarkastaja
Vesiensuojelu ja laki kestävän metsätalouden rahoituksesta (KEMERA) Jyväskylä 16.4.2013 Antti Leinonen Suomen metsäkeskus
 Jyväskylä 16.4.2013 Antti Leinonen Suomen metsäkeskus") Vesiensuojelu ja laki kestävän metsätalouden rahoituksesta (KEMERA) Jyväskylä 16.4.2013 Antti Leinonen Suomen metsäkeskus 1 Laki kestävän metsätalouden rahoituksesta (1994/1996) Laki (1994/1996) voimassa
Vesiensuojelu ja laki kestävän metsätalouden rahoituksesta (KEMERA) Jyväskylä 16.4.2013 Antti Leinonen Suomen metsäkeskus 1 Laki kestävän metsätalouden rahoituksesta (1994/1996) Laki (1994/1996) voimassa
Spatiaalisen sijainnin, ympäristötekijöiden ja lajien levittäytymiskyvyn vaikutukset pohjaeläinyhteisöjen rakenteeseen subarktisissa
 Spatiaalisen sijainnin, ympäristötekijöiden ja lajien levittäytymiskyvyn vaikutukset pohjaeläinyhteisöjen rakenteeseen subarktisissa virtavesissä Olli-Matti Kärnä Pro gradu -tutkielma 791619S Oulun yliopisto
Spatiaalisen sijainnin, ympäristötekijöiden ja lajien levittäytymiskyvyn vaikutukset pohjaeläinyhteisöjen rakenteeseen subarktisissa virtavesissä Olli-Matti Kärnä Pro gradu -tutkielma 791619S Oulun yliopisto
Schuelerin vettä läpäisemättömän pinnan osuuteen perustuvan taajamapurojen luokittelun soveltuvuus Vantaan pienvaluma-alueille
 Schuelerin vettä läpäisemättömän pinnan osuuteen perustuvan taajamapurojen luokittelun soveltuvuus Vantaan pienvaluma-alueille Outi Kesäniemi Taajamapuroja voidaan luokitella rakennetun pinta-alan perusteella
Schuelerin vettä läpäisemättömän pinnan osuuteen perustuvan taajamapurojen luokittelun soveltuvuus Vantaan pienvaluma-alueille Outi Kesäniemi Taajamapuroja voidaan luokitella rakennetun pinta-alan perusteella
Humusvedet. Tummien vesien ekologiaa. Lauri Arvola. Helsingin yliopisto Lammin biologinen asema
 Humusvedet Tummien vesien ekologiaa Lauri Arvola Helsingin yliopisto Lammin biologinen asema Sisältö Mitä humus on? Humusaineiden mittaamisesta Humusaineiden hajoaminen Mistä vesistöjen humusaineet ovat
Humusvedet Tummien vesien ekologiaa Lauri Arvola Helsingin yliopisto Lammin biologinen asema Sisältö Mitä humus on? Humusaineiden mittaamisesta Humusaineiden hajoaminen Mistä vesistöjen humusaineet ovat
Maaperän biologinen monimuotoisuus Tuhannet tuntemattomat jalkojemme alla
 Maaperän biologinen monimuotoisuus Tuhannet tuntemattomat jalkojemme alla Jari Haimi Bio- ja ympäristötieteiden laitos Jyväskylän yliopisto 24.11.2015 Maaperän monimuotoisuus 2 Maaperässä elää ja vaikuttaa
Maaperän biologinen monimuotoisuus Tuhannet tuntemattomat jalkojemme alla Jari Haimi Bio- ja ympäristötieteiden laitos Jyväskylän yliopisto 24.11.2015 Maaperän monimuotoisuus 2 Maaperässä elää ja vaikuttaa
Laboratorioanalyysit, vertailunäytteet ja tilastolliset menetelmät
 Jarmo Koskiniemi Maataloustieteiden laitos Helsingin yliopisto 0504151624 jarmo.koskiniemi@helsinki.fi 03.12.2015 Kolkunjoen taimenten geneettinen analyysi Näytteet Mika Oraluoma (Vesi-Visio osk) toimitti
Jarmo Koskiniemi Maataloustieteiden laitos Helsingin yliopisto 0504151624 jarmo.koskiniemi@helsinki.fi 03.12.2015 Kolkunjoen taimenten geneettinen analyysi Näytteet Mika Oraluoma (Vesi-Visio osk) toimitti
Bioteknologian perustyökaluja
 Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
Bioteknologian perustyökaluja DNAn ja RNAn eristäminen helppoa. Puhdistaminen työlästä (DNA pestään lukuisilla liuottimilla). Myös lähetti-rnat voidaan eristää ja muuntaa virusten käänteiskopioijaentsyymin
Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa
 Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa Maria Valkonen, Kaisa Jalkanen, Martin Täubel, Anne Hyvärinen 31.3.2014 Sisäilmastoseminaari 2014 1 Tausta Asumisterveysoppaan mukaiset sisäympäristön
Kvantitatiivisen PCR:n käyttö mikrobivaurion toteamisessa Maria Valkonen, Kaisa Jalkanen, Martin Täubel, Anne Hyvärinen 31.3.2014 Sisäilmastoseminaari 2014 1 Tausta Asumisterveysoppaan mukaiset sisäympäristön
Riekon (Lagopus lagopus) talviravinnon käyttö ja valinta Suomen eteläisissä populaatioissa
 talviravinnon käyttö ja valinta Suomen eteläisissä populaatioissa") Riekon (Lagopus lagopus) talviravinnon käyttö ja valinta Suomen eteläisissä populaatioissa Jenni Miettunen Pro gradu 2009 Ohjaaja: Prof. Heikki Roininen Biotieteiden tiedekunta Joensuun yliopisto Riekon
Riekon (Lagopus lagopus) talviravinnon käyttö ja valinta Suomen eteläisissä populaatioissa Jenni Miettunen Pro gradu 2009 Ohjaaja: Prof. Heikki Roininen Biotieteiden tiedekunta Joensuun yliopisto Riekon
Biologia. Pakolliset kurssit. 1. Eliömaailma (BI1)
") Biologia Pakolliset kurssit 1. Eliömaailma (BI1) tuntee elämän tunnusmerkit ja perusedellytykset sekä tietää, miten elämän ilmiöitä tutkitaan ymmärtää, mitä luonnon monimuotoisuus biosysteemien eri tasoilla
Biologia Pakolliset kurssit 1. Eliömaailma (BI1) tuntee elämän tunnusmerkit ja perusedellytykset sekä tietää, miten elämän ilmiöitä tutkitaan ymmärtää, mitä luonnon monimuotoisuus biosysteemien eri tasoilla
Mitä pajukosteikon juuriston mikrobianalyysi kertoo? , Oulun yliopisto Karoliina Iso-Junno
 Mitä pajukosteikon juuriston mikrobianalyysi kertoo?, Oulun yliopisto Karoliina Iso-Junno Sisältö Taustaa Työn tarkoitus Materiaalit ja menetelmät Tulokset&Pohdinta Yhteenveto Taustaa - Mikrobit Mikro-organismit:
Mitä pajukosteikon juuriston mikrobianalyysi kertoo?, Oulun yliopisto Karoliina Iso-Junno Sisältö Taustaa Työn tarkoitus Materiaalit ja menetelmät Tulokset&Pohdinta Yhteenveto Taustaa - Mikrobit Mikro-organismit:
LOHJAN JÄRVIEN VEDENLAATUSEURANTA 2012 Kaitalampi
 LUVY/109 27.7.2012 Risto Murto Lohjan kaupunki ympäristönsuojelu LOHJAN JÄRVIEN VEDENLAATUSEURANTA 2012 Kaitalampi Näytteenotto liittyy Lohjan kaupungin lakisääteiseen velvoitteeseen seurata ympäristön
LUVY/109 27.7.2012 Risto Murto Lohjan kaupunki ympäristönsuojelu LOHJAN JÄRVIEN VEDENLAATUSEURANTA 2012 Kaitalampi Näytteenotto liittyy Lohjan kaupungin lakisääteiseen velvoitteeseen seurata ympäristön
LUONNONHUUHTOUMA Tietoa luonnonhuuhtoumasta tarvitaan ihmisen aiheuttaman kuormituksen arvioimiseksi Erityisesti metsätalous
 LUONNONHUUHTOUMA Tietoa luonnonhuuhtoumasta tarvitaan ihmisen aiheuttaman kuormituksen arvioimiseksi Erityisesti metsätalous Luonnonhuuhtoumaan vaikuttavat mm.: Geologia, ilmasto Maaperä, topografia, kasvillisuus
LUONNONHUUHTOUMA Tietoa luonnonhuuhtoumasta tarvitaan ihmisen aiheuttaman kuormituksen arvioimiseksi Erityisesti metsätalous Luonnonhuuhtoumaan vaikuttavat mm.: Geologia, ilmasto Maaperä, topografia, kasvillisuus
POHJAVEDEN IN SITU PUHDISTAMINEN UUDELLA MENETELMÄSOVELLUKSELLA
 POHJAVEDEN IN SITU PUHDISTAMINEN UUDELLA MENETELMÄSOVELLUKSELLA CASE KÄRKÖLÄ KLOORIFENOLEILLA PILAANTUNUT POHJAVESIALUE JUKKA IKÄHEIMO, PÖYRY FINLAND OY II IV Jukka Ikäheimo 19.3.2015 1 KOKEET KLOORIFENOLIEN
POHJAVEDEN IN SITU PUHDISTAMINEN UUDELLA MENETELMÄSOVELLUKSELLA CASE KÄRKÖLÄ KLOORIFENOLEILLA PILAANTUNUT POHJAVESIALUE JUKKA IKÄHEIMO, PÖYRY FINLAND OY II IV Jukka Ikäheimo 19.3.2015 1 KOKEET KLOORIFENOLIEN
MERIKARVIA. Merikarviantien alkupään ja Yrittäjäntien ympäristön asemakaavoitus. Hulevesitarkastelu. Kankaanpään kaupunki. Ympäristökeskus.
 Hulevesitarkastelu Kankaanpään kaupunki Ympäristökeskus talvi 2015 v.2 SISÄLLYS Hulevesien hallinta 2 Kaavoitettavan alueen sijainti 2 Valuma-alue 3 Hulevedet kaava-alueella 4 Hulevesimäärät 5-6 1 HULEVESIEN
Hulevesitarkastelu Kankaanpään kaupunki Ympäristökeskus talvi 2015 v.2 SISÄLLYS Hulevesien hallinta 2 Kaavoitettavan alueen sijainti 2 Valuma-alue 3 Hulevedet kaava-alueella 4 Hulevesimäärät 5-6 1 HULEVESIEN
Vesiensuojelutoimenpiteiden vaikutusten mittaaminen vesistössä. Pasi Valkama Vantaanjoen ja Helsingin seudun vesiensuojeluyhdistys ry
 Vesiensuojelutoimenpiteiden vaikutusten mittaaminen vesistössä Vantaanjoen ja Helsingin seudun vesiensuojeluyhdistys ry Esityksen sisältö Lyhyesti automaattisesta veden laadun seurannasta Kasvipeite/muokkaus/
Vesiensuojelutoimenpiteiden vaikutusten mittaaminen vesistössä Vantaanjoen ja Helsingin seudun vesiensuojeluyhdistys ry Esityksen sisältö Lyhyesti automaattisesta veden laadun seurannasta Kasvipeite/muokkaus/
Vesiensuojelu metsänuudistamisessa kivennäismailla
 Vesiensuojelu metsänuudistamisessa kivennäismailla Sirpa Piirainen, MMT, varttunut tutkija Esitykseni sisältö Taustatietoja Pohjavedet ja metsänuudistaminen Huuhtoumat vesistöihin Vesiensuojelun avainkohdat
Vesiensuojelu metsänuudistamisessa kivennäismailla Sirpa Piirainen, MMT, varttunut tutkija Esitykseni sisältö Taustatietoja Pohjavedet ja metsänuudistaminen Huuhtoumat vesistöihin Vesiensuojelun avainkohdat
Yleiskatsaus metsätalouden vesistövaikutuksiin ja vesiensuojelun lainsäädäntöön
 Yleiskatsaus metsätalouden vesistövaikutuksiin ja vesiensuojelun lainsäädäntöön Samuli Joensuu Lapua 12.11.2013 Sisältö Metsätalouden kuormitusvaikutuksista Muuttuva lainsäädäntö ja sen merkitys metsätalouden
Yleiskatsaus metsätalouden vesistövaikutuksiin ja vesiensuojelun lainsäädäntöön Samuli Joensuu Lapua 12.11.2013 Sisältö Metsätalouden kuormitusvaikutuksista Muuttuva lainsäädäntö ja sen merkitys metsätalouden
UIMAVESIPROFIILI HUUTJÄRVEN UIMARANTA
 Pyhtään kunta 20.4.2016 UIMAVESIPROFIILI HUUTJÄRVEN UIMARANTA 1. YHTEYSTIEDOT 1.1 Uimarannan omistaja ja Pyhtään kunta, Siltakyläntie 175, 49220 Siltakylä 1.2 Uimarannan päävastuullinen hoitaja ja 1.3
Pyhtään kunta 20.4.2016 UIMAVESIPROFIILI HUUTJÄRVEN UIMARANTA 1. YHTEYSTIEDOT 1.1 Uimarannan omistaja ja Pyhtään kunta, Siltakyläntie 175, 49220 Siltakylä 1.2 Uimarannan päävastuullinen hoitaja ja 1.3
BIOMUOVIA TÄRKKELYKSESTÄ
 BIOMUOVIA TÄRKKELYKSESTÄ KOHDERYHMÄ: Soveltuu peruskoulun 9.luokan kemian osioon Orgaaninen kemia. KESTO: 45 60 min. Kemian opetuksen keskus MOTIVAATIO: Muovituotteet kerääntyvät helposti luontoon ja saastuttavat
BIOMUOVIA TÄRKKELYKSESTÄ KOHDERYHMÄ: Soveltuu peruskoulun 9.luokan kemian osioon Orgaaninen kemia. KESTO: 45 60 min. Kemian opetuksen keskus MOTIVAATIO: Muovituotteet kerääntyvät helposti luontoon ja saastuttavat
Miten arvokkaat pienvedet tunnistetaan maastossa? Metsätalouden vesiensuojelupäivät, Koli Jari Ilmonen, Luontopalvelut
 Miten arvokkaat pienvedet tunnistetaan maastossa? 22.09.2015 Metsätalouden vesiensuojelupäivät, Koli Jari Ilmonen, Luontopalvelut Mitä ovat arvokkaat pienvedet? Pienvedet = purot ja norot, lammet, lähteiköt
Miten arvokkaat pienvedet tunnistetaan maastossa? 22.09.2015 Metsätalouden vesiensuojelupäivät, Koli Jari Ilmonen, Luontopalvelut Mitä ovat arvokkaat pienvedet? Pienvedet = purot ja norot, lammet, lähteiköt
3 MALLASVEDEN PINNAN KORKEUS
 1 TAVASE OY, IMEYTYS- JA MERKKIAINEKOKEEN AIKAISEN TARKKAILUN YHTEENVETO 26.4.2010 1 YLEISTÄ Tavase Oy toteuttaa tekopohjavesihankkeen imeytys- ja merkkiainekokeen tutkimusalueellaan Syrjänharjussa Pälkäneellä.
1 TAVASE OY, IMEYTYS- JA MERKKIAINEKOKEEN AIKAISEN TARKKAILUN YHTEENVETO 26.4.2010 1 YLEISTÄ Tavase Oy toteuttaa tekopohjavesihankkeen imeytys- ja merkkiainekokeen tutkimusalueellaan Syrjänharjussa Pälkäneellä.
VIRTAVESIEN KUNNOSTUS. Pauliina Louhi Kalaekologian tutkimusseminaari
 VIRTAVESIEN KUNNOSTUS Pauliina Louhi Kalaekologian tutkimusseminaari 10.4.2012 Sisältö 1. Kunnostuksien ekologinen tausta ja tavoitteet 2. Uittoperkaukset Suomessa 3. Virtavesien kunnostukset Suomessa
VIRTAVESIEN KUNNOSTUS Pauliina Louhi Kalaekologian tutkimusseminaari 10.4.2012 Sisältö 1. Kunnostuksien ekologinen tausta ja tavoitteet 2. Uittoperkaukset Suomessa 3. Virtavesien kunnostukset Suomessa
NIINIMÄEN TUULIPUISTO OY Sähkönsiirtolinjojen liito-oravaselvitys, Pieksämäki
 RAPORTTI 16X267156_E722 13.4.2016 NIINIMÄEN TUULIPUISTO OY Sähkönsiirtolinjojen liito-oravaselvitys, Pieksämäki 1 Niinimäen Tuulipuisto Oy Sähkönsiirtolinjojen liito-oravaselvitys, Pieksämäki Sisältö 1
RAPORTTI 16X267156_E722 13.4.2016 NIINIMÄEN TUULIPUISTO OY Sähkönsiirtolinjojen liito-oravaselvitys, Pieksämäki 1 Niinimäen Tuulipuisto Oy Sähkönsiirtolinjojen liito-oravaselvitys, Pieksämäki Sisältö 1
UIMAVESIPROFIILI RANTAKYLÄN UIMAPAIKKA
 3.12.2012 UIMAVESIPROFIILI RANTAKYLÄN UIMAPAIKKA SISÄLLYS 1. YHTEYSTIEDOT 1.1 Uimarannan omistaja ja yhteystiedot 1.2 Uimarannan päävastuullinen hoitaja ja yhteystiedot 1.3 Uimarantaa valvova viranomainen
3.12.2012 UIMAVESIPROFIILI RANTAKYLÄN UIMAPAIKKA SISÄLLYS 1. YHTEYSTIEDOT 1.1 Uimarannan omistaja ja yhteystiedot 1.2 Uimarannan päävastuullinen hoitaja ja yhteystiedot 1.3 Uimarantaa valvova viranomainen
ristöjen hoito - Vesilinnut
 Elinympärist ristöjen hoito - Vesilinnut Vesilintuelinympärist ristöt t = vesiensuojelu + maisema + luonnon Piirrokset: Jari Kostet ja MKJ Kuvat: Mikko Alhainen, Marko Svensberg, Marko Muuttola, Harri
Elinympärist ristöjen hoito - Vesilinnut Vesilintuelinympärist ristöt t = vesiensuojelu + maisema + luonnon Piirrokset: Jari Kostet ja MKJ Kuvat: Mikko Alhainen, Marko Svensberg, Marko Muuttola, Harri
Purojen, järvien ja jokien kemiallisen, fysikaalisen ja biologisen tilan muutokset ja niiden merkitys
 Purojen, järvien ja jokien kemiallisen, fysikaalisen ja biologisen tilan muutokset ja niiden merkitys Kari-Matti Vuori & Mika Visuri Metsätalous ja vesistöt seminaari 26.9. 2006, Koli SISÄLTÖ TILAA MITTAAVAT
Purojen, järvien ja jokien kemiallisen, fysikaalisen ja biologisen tilan muutokset ja niiden merkitys Kari-Matti Vuori & Mika Visuri Metsätalous ja vesistöt seminaari 26.9. 2006, Koli SISÄLTÖ TILAA MITTAAVAT
Humuksen vaikutukset järvien hiilenkiertoon ja ravintoverkostoihin. Paula Kankaala FT, dos. Itä Suomen yliopisto Biologian laitos
 Humuksen vaikutukset järvien hiilenkiertoon ja ravintoverkostoihin Paula Kankaala FT, dos. Itä Suomen yliopisto Biologian laitos Hiilenkierto järvessä Valuma alueelta peräisin oleva orgaaninen aine (humus)
Humuksen vaikutukset järvien hiilenkiertoon ja ravintoverkostoihin Paula Kankaala FT, dos. Itä Suomen yliopisto Biologian laitos Hiilenkierto järvessä Valuma alueelta peräisin oleva orgaaninen aine (humus)
Suomen huonosti tunnetut ja uhanalaiset sienet
 Suomen huonosti tunnetut ja uhanalaiset sienet Sienet Sienten lajimäärä on paljon aiemmin ajateltua suurempi Lajit tunnettava, jotta sienten todellisen monimuotoisuuden, uhanalaisuuden ja suojelutarpeiden
Suomen huonosti tunnetut ja uhanalaiset sienet Sienet Sienten lajimäärä on paljon aiemmin ajateltua suurempi Lajit tunnettava, jotta sienten todellisen monimuotoisuuden, uhanalaisuuden ja suojelutarpeiden
VALKJÄRVEN VEDEN LAATU Kesän 2015 tutkimus ja vertailu kesiin 2010-2014
 LUVY/121 6.7.215 Anne Linnonmaa Valkjärven suojeluyhdistys ry anne.linnonmaa@anne.fi VALKJÄRVEN VEDEN LAATU Kesän 215 tutkimus ja vertailu kesiin 21-214 Sammatin Valkjärvestä otettiin vesinäytteet 25.6.215
LUVY/121 6.7.215 Anne Linnonmaa Valkjärven suojeluyhdistys ry anne.linnonmaa@anne.fi VALKJÄRVEN VEDEN LAATU Kesän 215 tutkimus ja vertailu kesiin 21-214 Sammatin Valkjärvestä otettiin vesinäytteet 25.6.215
Luonnonmukainen vesirakentaminen peruskuivatushankkeissa. Lasse Järvenpää, SYKE Salaojateknikoiden neuvottelupäivät, 1.2.
 Luonnonmukainen vesirakentaminen peruskuivatushankkeissa Lasse Järvenpää, SYKE Salaojateknikoiden neuvottelupäivät, 1.2.2007, Hyvinkää Esityksen aiheet Perattujen purojen kunnostus ja hoito Monitavoitteiset
Luonnonmukainen vesirakentaminen peruskuivatushankkeissa Lasse Järvenpää, SYKE Salaojateknikoiden neuvottelupäivät, 1.2.2007, Hyvinkää Esityksen aiheet Perattujen purojen kunnostus ja hoito Monitavoitteiset
Paimion Karhunojan vedenlaatututkimukset vuonna 2015
 1(4) 16.12.2015 Paimion Karhunojan vedenlaatututkimukset vuonna 2015 1 YLEISTÄ Lounais-Suomen vesiensuojeluyhdistys ry tutki Paimion Karhunojan vedenlaatua vuonna 2015 jatkuvatoimisella MS5 Hydrolab vedenlaatumittarilla
1(4) 16.12.2015 Paimion Karhunojan vedenlaatututkimukset vuonna 2015 1 YLEISTÄ Lounais-Suomen vesiensuojeluyhdistys ry tutki Paimion Karhunojan vedenlaatua vuonna 2015 jatkuvatoimisella MS5 Hydrolab vedenlaatumittarilla
Suomen vesistöjen tummuminen. Antti Räike Suomen ympäristökeskus Merikeskus
 Suomen vesistöjen tummuminen Antti Räike Suomen ympäristökeskus Merikeskus Mitä vesien tummumisella tarkoitetaan? Kuva: Stefan Löfgren Tummumisella käsitetään humuksen lisääntymistä, joka ilmenee veden
Suomen vesistöjen tummuminen Antti Räike Suomen ympäristökeskus Merikeskus Mitä vesien tummumisella tarkoitetaan? Kuva: Stefan Löfgren Tummumisella käsitetään humuksen lisääntymistä, joka ilmenee veden
Valuma-alueen merkitys vesiensuojelussa
 Valuma-alueen merkitys vesiensuojelussa Marjo Tarvainen Asiantuntija, FT Pyhäjärvi-instituutti 25.1.2010 VOPPE koulutus, Eura 1 Veden laatuun vaikuttavia tekijöitä Vesitase Sateet lisäävät virtaamia, mitkä
Valuma-alueen merkitys vesiensuojelussa Marjo Tarvainen Asiantuntija, FT Pyhäjärvi-instituutti 25.1.2010 VOPPE koulutus, Eura 1 Veden laatuun vaikuttavia tekijöitä Vesitase Sateet lisäävät virtaamia, mitkä
Rehevöityneen järven kunnostamisen haasteet
 Rehevöityneen järven kunnostamisen haasteet , N Hyväkuntoinen terve järvi kestää ravinnekuormitusta varsin hyvin ilman, että veden laatu suuresti muuttuu, koska lukuisat puskurimekanismit ehkäisevät muutosta
Rehevöityneen järven kunnostamisen haasteet , N Hyväkuntoinen terve järvi kestää ravinnekuormitusta varsin hyvin ilman, että veden laatu suuresti muuttuu, koska lukuisat puskurimekanismit ehkäisevät muutosta
Virtavesikunnostusten vaikutukset jokiluonnon ja ekosysteemipalvelujen näkökulmasta
 Virtavesikunnostusten vaikutukset jokiluonnon ja ekosysteemipalvelujen näkökulmasta Maare Marttila Vesistökunnostusverkoston vuosiseminaari 2017 Elinympäristökunnostukset Ensimmäiset kunnostukset 1970-luvulla,
Virtavesikunnostusten vaikutukset jokiluonnon ja ekosysteemipalvelujen näkökulmasta Maare Marttila Vesistökunnostusverkoston vuosiseminaari 2017 Elinympäristökunnostukset Ensimmäiset kunnostukset 1970-luvulla,
Johtuuko tämä ilmastonmuutoksesta? - kasvihuoneilmiön voimistuminen vaikutus sääolojen vaihteluun
 Johtuuko tämä ilmastonmuutoksesta? - kasvihuoneilmiön voimistuminen vaikutus sääolojen vaihteluun Jouni Räisänen Helsingin yliopiston fysiikan laitos 15.1.2010 Vuorokauden keskilämpötila Talvi 2007-2008
Johtuuko tämä ilmastonmuutoksesta? - kasvihuoneilmiön voimistuminen vaikutus sääolojen vaihteluun Jouni Räisänen Helsingin yliopiston fysiikan laitos 15.1.2010 Vuorokauden keskilämpötila Talvi 2007-2008
Hämeenlinnan ja Janakkalan Valajärven tila. Heli Jutila ympäristötarkastaja 1.6.2009
 Hämeenlinnan ja Janakkalan Valajärven tila Heli Jutila ympäristötarkastaja 1.6.2009 Valajärven valuma-alue Soita, metsää, harjuja; vähän peltoja: 15,01 km 2 : 4,3 x järven ala eli ei erityisen suuri 2.6.2009
Hämeenlinnan ja Janakkalan Valajärven tila Heli Jutila ympäristötarkastaja 1.6.2009 Valajärven valuma-alue Soita, metsää, harjuja; vähän peltoja: 15,01 km 2 : 4,3 x järven ala eli ei erityisen suuri 2.6.2009
Harjuskannan tila ja luonnonvaraisen lisääntymisen mahdollisuudet Kokemäenjoessa
 Harjuskannan tila ja luonnonvaraisen lisääntymisen mahdollisuudet Kokemäenjoessa Tutkimusalue Kokemäenjoessa TEHDYT TUTKIMUKSET Nykytilan selvittäminen: - kalastustiedustelu - perhokalastajien haastattelu
Harjuskannan tila ja luonnonvaraisen lisääntymisen mahdollisuudet Kokemäenjoessa Tutkimusalue Kokemäenjoessa TEHDYT TUTKIMUKSET Nykytilan selvittäminen: - kalastustiedustelu - perhokalastajien haastattelu
Taustaa puustoisista perinneympäristöistä
 Taustaa puustoisista perinneympäristöistä Laitila 4.- 5.9.2012 Hannele Kekäläinen ylitarkastaja Etelä-Pohjanmaan ELY-keskus, Ympäristö- ja luonnonvarat vastuualue Maatalousympäristöt Suomen viidenneksi
Taustaa puustoisista perinneympäristöistä Laitila 4.- 5.9.2012 Hannele Kekäläinen ylitarkastaja Etelä-Pohjanmaan ELY-keskus, Ympäristö- ja luonnonvarat vastuualue Maatalousympäristöt Suomen viidenneksi
Purokunnostuksia Iijoen vesistössä Koillismaalla. Pirkko-Liisa Luhta, Eero Moilanen, Matti Suanto Luontopalvelut
 Purokunnostuksia Iijoen vesistössä Koillismaalla Pirkko-Liisa Luhta, Eero Moilanen, Matti Suanto Luontopalvelut Esityksen sisältö Purojen inventointi Kunnostukset menetelmineen Vaikutusten seuranta Mitä
Purokunnostuksia Iijoen vesistössä Koillismaalla Pirkko-Liisa Luhta, Eero Moilanen, Matti Suanto Luontopalvelut Esityksen sisältö Purojen inventointi Kunnostukset menetelmineen Vaikutusten seuranta Mitä
Valuma-alueen merkitys vesien tilan parantamisessa. Vanajavesikeskus-hankkeen Vesistöasiantuntija Suvi Mäkelä
 Valuma-alueen merkitys vesien tilan parantamisessa Vanajavesikeskus-hankkeen Vesistöasiantuntija Suvi Mäkelä Mikä valuma-alue? Kuinka kauas pitää katsoa? Lähivaluma-alue Kaukovaluma-alue Latvavedet 2.
Valuma-alueen merkitys vesien tilan parantamisessa Vanajavesikeskus-hankkeen Vesistöasiantuntija Suvi Mäkelä Mikä valuma-alue? Kuinka kauas pitää katsoa? Lähivaluma-alue Kaukovaluma-alue Latvavedet 2.
Rantamo-Seittelin kosteikon vedenlaadun seuranta
 Rantamo-Seittelin kosteikon vedenlaadun seuranta Jari Koskiaho, SYKE Tuusulanjärven tila paremmaksi -seminaari Gustavelund 23.5.2013 Kosteikoissa tapahtuvat vedenpuhdistusprosessit Kiintoaineksen laskeutuminen
Rantamo-Seittelin kosteikon vedenlaadun seuranta Jari Koskiaho, SYKE Tuusulanjärven tila paremmaksi -seminaari Gustavelund 23.5.2013 Kosteikoissa tapahtuvat vedenpuhdistusprosessit Kiintoaineksen laskeutuminen
TAVASE OY, IMEYTYS- JA MERKKIAINEKOKEEN AIKAISEN TARKKAILUN YHTEENVETO 24.6.2010
 1 TAVASE OY, IMEYTYS- JA MERKKIAINEKOKEEN AIKAISEN TARKKAILUN YHTEENVETO 24.6.2010 1 YLEISTÄ Tavase Oy toteuttaa tekopohjavesihankkeen imeytys- ja merkkiainekokeen tutkimusalueellaan Syrjänharjussa Pälkäneellä.
1 TAVASE OY, IMEYTYS- JA MERKKIAINEKOKEEN AIKAISEN TARKKAILUN YHTEENVETO 24.6.2010 1 YLEISTÄ Tavase Oy toteuttaa tekopohjavesihankkeen imeytys- ja merkkiainekokeen tutkimusalueellaan Syrjänharjussa Pälkäneellä.
Vesilaki 2011 ja metsäojitukset
 Vesilaki 2011 ja metsäojitukset Puron ja noron luonnontilan turvaaminen Vähäinen ojitus Ilmoitusmenettelystä Pekka Kemppainen Maa- ja metsätalousministeriö Metsätalouden vesiensuojelupäivät Saarijärvi
Vesilaki 2011 ja metsäojitukset Puron ja noron luonnontilan turvaaminen Vähäinen ojitus Ilmoitusmenettelystä Pekka Kemppainen Maa- ja metsätalousministeriö Metsätalouden vesiensuojelupäivät Saarijärvi
Liito-orava kartoitus Nouvanlahden ulkoilualueelle sekä eteläisen Kilpijärven länsirannalle.
 Liito-orava kartoitus Nouvanlahden ulkoilualueelle sekä eteläisen Kilpijärven länsirannalle. Tarmo Saastamoinen 2010. Kuva.1 Kaatunut kuusenrunko Nouvanlahdesta. LIITO-ORAVA: Liito-orava (pteromys volans)on
Liito-orava kartoitus Nouvanlahden ulkoilualueelle sekä eteläisen Kilpijärven länsirannalle. Tarmo Saastamoinen 2010. Kuva.1 Kaatunut kuusenrunko Nouvanlahdesta. LIITO-ORAVA: Liito-orava (pteromys volans)on
PUROT JA OJAT LÄHILUONNOSSA
 PUROT JA OJAT LÄHILUONNOSSA Somero 26.9.2014 Matti Jantunen Lounais-Suomen vesiensuojeluyhdistys ry Mikä on puro? Vesilaki (27.5.2011/587) 3 määrittelee: Joki = virtaavan veden vesistö, jonka valuma-alue
PUROT JA OJAT LÄHILUONNOSSA Somero 26.9.2014 Matti Jantunen Lounais-Suomen vesiensuojeluyhdistys ry Mikä on puro? Vesilaki (27.5.2011/587) 3 määrittelee: Joki = virtaavan veden vesistö, jonka valuma-alue
KORPI Bioenergiakorjuun ekologiset vesistövaikutukset
 KORPI Bioenergiakorjuun ekologiset vesistövaikutukset Muokkaa alaotsikon perustyyliä naps. Vesiensuojelu turvemailla seminaari, 8.4.2011, Seinäjoki FT Kristian Meissner, SYKE FM Mika Nieminen, Jyväskylän
KORPI Bioenergiakorjuun ekologiset vesistövaikutukset Muokkaa alaotsikon perustyyliä naps. Vesiensuojelu turvemailla seminaari, 8.4.2011, Seinäjoki FT Kristian Meissner, SYKE FM Mika Nieminen, Jyväskylän
Tutkimusmateriaalit -ja välineet: kaarnan palaset, hiekan murut, pihlajanmarjat, juuripalat, pakasterasioita, vettä, suolaa ja porkkananpaloja.
 JIPPO-POLKU Jippo-polku sisältää kokeellisia tutkimustehtäviä toteutettavaksi perusopetuksessa, kerhossa tai kotona. Polun tehtävät on tarkoitettu suoritettavaksi luonnossa joko koulun tai kerhon lähimaastossa,
JIPPO-POLKU Jippo-polku sisältää kokeellisia tutkimustehtäviä toteutettavaksi perusopetuksessa, kerhossa tai kotona. Polun tehtävät on tarkoitettu suoritettavaksi luonnossa joko koulun tai kerhon lähimaastossa,
Retkiä Mynälahdelle retkiesimerkit
 Retkiä Mynälahdelle retkiesimerkit Sisällysluettelo Alkusanat... 1 Puuretki pienimmille oppilaille... 2 Halataan puita!... 2 Erilaiset puut... 2 Elämää vanhoissa puissa... 3 Lempipuu... 3 Vesistöretki
Retkiä Mynälahdelle retkiesimerkit Sisällysluettelo Alkusanat... 1 Puuretki pienimmille oppilaille... 2 Halataan puita!... 2 Erilaiset puut... 2 Elämää vanhoissa puissa... 3 Lempipuu... 3 Vesistöretki
29.03.2006 RATU rankkasateet ja taajamatulvat TKK:n vesitalouden ja vesirakennuksen hankeosien tilanne ja välitulokset T. Karvonen ja T.
 29.3.26 RATU rankkasateet ja taajamatulvat TKK:n vesitalouden ja vesirakennuksen hankeosien tilanne ja välitulokset T. Karvonen ja T. Tiihonen RATU/TKK:n osuus Laaditaan kahdentyyppisiä malleja: * taajamavesien
29.3.26 RATU rankkasateet ja taajamatulvat TKK:n vesitalouden ja vesirakennuksen hankeosien tilanne ja välitulokset T. Karvonen ja T. Tiihonen RATU/TKK:n osuus Laaditaan kahdentyyppisiä malleja: * taajamavesien
Lupahakemuksen täydennys
 Lupahakemuksen täydennys 26.4.2012 Talvivaara Sotkamo Oy Talvivaarantie 66 88120 Tuhkakylä Finland 2012-04-26 2 / 6 Lupahakemuksen täydennys Täydennyskehotuksessa (11.4.2012) täsmennettäväksi pyydetyt
Lupahakemuksen täydennys 26.4.2012 Talvivaara Sotkamo Oy Talvivaarantie 66 88120 Tuhkakylä Finland 2012-04-26 2 / 6 Lupahakemuksen täydennys Täydennyskehotuksessa (11.4.2012) täsmennettäväksi pyydetyt
Liuenneen hiilen (CDOM) laatu menetelmän soveltaminen turv le. Jonna Kuha, Toni Roiha, Mika Nieminen,Hannu Marttila
 laatu menetelmän soveltaminen turv le. Jonna Kuha, Toni Roiha, Mika Nieminen,Hannu Marttila") Liuenneen hiilen (CDOM) laatu menetelmän soveltaminen turvemaille Jonna Kuha, Toni Roiha, Mika Nieminen,Hannu Marttila Mitä humusaineet ovat? Liuenneen eloperäisen (orgaanisen) aineksen eli humuksen värillinen
Liuenneen hiilen (CDOM) laatu menetelmän soveltaminen turvemaille Jonna Kuha, Toni Roiha, Mika Nieminen,Hannu Marttila Mitä humusaineet ovat? Liuenneen eloperäisen (orgaanisen) aineksen eli humuksen värillinen
Automaattimittarit valuma-alueella tehtävien kunnostustoimien vaikutusten seurannassa
 Automaattimittarit valuma-alueella tehtävien kunnostustoimien vaikutusten seurannassa Vantaanjoen ja Helsingin seudun vesiensuojeluyhdistys ry 14.6.2017 Esityksen sisältö Miksi automaattimittauksia kannattaa
Automaattimittarit valuma-alueella tehtävien kunnostustoimien vaikutusten seurannassa Vantaanjoen ja Helsingin seudun vesiensuojeluyhdistys ry 14.6.2017 Esityksen sisältö Miksi automaattimittauksia kannattaa
Puruvesi-seminaari 26.7.2014. Vastuunjako ja yhteistoiminnan järjestelyt vesiensuojelussa. Ylijohtaja Pekka Häkkinen Etelä-Savon ELY-keskus
 Puruvesi-seminaari 26.7.2014 Vastuunjako ja yhteistoiminnan järjestelyt vesiensuojelussa Ylijohtaja Pekka Häkkinen Etelä-Savon ELY-keskus Suomen pintavesien ekologinen tilaluokitus julkaistiin 2.10.2013.
Puruvesi-seminaari 26.7.2014 Vastuunjako ja yhteistoiminnan järjestelyt vesiensuojelussa Ylijohtaja Pekka Häkkinen Etelä-Savon ELY-keskus Suomen pintavesien ekologinen tilaluokitus julkaistiin 2.10.2013.
JA MUITA MENETELMIÄ PILAANTUNEIDEN SEDIMENTTIEN KÄSITTELYYN. Päivi Seppänen, Golder Associates Oy
 GEOTEKSTIILIALLAS JA MUITA MENETELMIÄ PILAANTUNEIDEN SEDIMENTTIEN KÄSITTELYYN Päivi Seppänen, Golder Associates Oy Käsittelymenetelmät ESITYKSEN RAKENNE Vedenpoistomenetelmät Puhdistusmenetelmät Sijoitusmenetelmät
GEOTEKSTIILIALLAS JA MUITA MENETELMIÄ PILAANTUNEIDEN SEDIMENTTIEN KÄSITTELYYN Päivi Seppänen, Golder Associates Oy Käsittelymenetelmät ESITYKSEN RAKENNE Vedenpoistomenetelmät Puhdistusmenetelmät Sijoitusmenetelmät
Kuinka vesipuitedirektiivi ja muu ympäristölainsäädäntö ohjaa metsätalouden vesiensuojelua
 Kuinka vesipuitedirektiivi ja muu ympäristölainsäädäntö ohjaa metsätalouden vesiensuojelua Paula Mononen Pohjois-Karjalan ympäristökeskus Metsätalous ja vesistöt -seminaari, Koli 26.9.2006 Vesipolitiikan
Kuinka vesipuitedirektiivi ja muu ympäristölainsäädäntö ohjaa metsätalouden vesiensuojelua Paula Mononen Pohjois-Karjalan ympäristökeskus Metsätalous ja vesistöt -seminaari, Koli 26.9.2006 Vesipolitiikan
MAAPERÄN HAJOTTAJAELIÖSTÖ MONTA-tutkimuksessa
 MAAPERÄN HAJOTTAJAELIÖSTÖ MONTA-tutkimuksessa Jari Haimi Jyväskylän yliopisto, Bio- ja ympäristötieteiden laitos Jyväskylän yliopisto Metsäntutkimuslaitos Metsäteho Tutkimuksessa ovat mukana olleeet: Anne
MAAPERÄN HAJOTTAJAELIÖSTÖ MONTA-tutkimuksessa Jari Haimi Jyväskylän yliopisto, Bio- ja ympäristötieteiden laitos Jyväskylän yliopisto Metsäntutkimuslaitos Metsäteho Tutkimuksessa ovat mukana olleeet: Anne
Vesilintujen runsauden muutoksia seurantaa, syitä. Jukka Kauppinen 2010
 Vesilintujen runsauden muutoksia seurantaa, syitä Jukka Kauppinen 2010 Vesilinnuston pitkäaikaisseurantaa ik i Pohjois-Savon järvien linnustokehityksestä on seuranta-aineistoa 1930-luvun puolivälistä alkaen.
Vesilintujen runsauden muutoksia seurantaa, syitä Jukka Kauppinen 2010 Vesilinnuston pitkäaikaisseurantaa ik i Pohjois-Savon järvien linnustokehityksestä on seuranta-aineistoa 1930-luvun puolivälistä alkaen.
Metsäpatologian laboratorio tuhotutkimuksen apuna. Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari
 Metsäpatologian laboratorio tuhotutkimuksen apuna Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari Metsäpuiden vaivat Metsäpuiden eloa ja terveyttä uhkaavat monet taudinaiheuttajat: Bioottiset taudinaiheuttajat
Metsäpatologian laboratorio tuhotutkimuksen apuna Metsätaimitarhapäivät 23. 24.1.2014 Anne Uimari Metsäpuiden vaivat Metsäpuiden eloa ja terveyttä uhkaavat monet taudinaiheuttajat: Bioottiset taudinaiheuttajat
Lääkeainejäämät biokaasulaitosten lopputuotteissa. Marja Lehto, MTT
 Kestävästi Kiertoon - seminaari Lääkeainejäämät biokaasulaitosten lopputuotteissa Marja Lehto, MTT Orgaaniset haitta-aineet aineet Termillä tarkoitetaan erityyppisiä orgaanisia aineita, joilla on jokin
Kestävästi Kiertoon - seminaari Lääkeainejäämät biokaasulaitosten lopputuotteissa Marja Lehto, MTT Orgaaniset haitta-aineet aineet Termillä tarkoitetaan erityyppisiä orgaanisia aineita, joilla on jokin
Turvepeltojen ympäristöhaasteet
 Turvepeltojen ympäristöhaasteet Kristiina Regina Turvepeltojen parhaat viljelytavat nyt ja tulevaisuudessa Ilmajoki 21.11.2017 Turvemaiden globaali merkitys Peittävät 3 % maa-alasta Varastoivat 30 % maaperän
Turvepeltojen ympäristöhaasteet Kristiina Regina Turvepeltojen parhaat viljelytavat nyt ja tulevaisuudessa Ilmajoki 21.11.2017 Turvemaiden globaali merkitys Peittävät 3 % maa-alasta Varastoivat 30 % maaperän
Kunnostusten seuranta ja seurantatutkimukset
 Kunnostusten seuranta ja seurantatutkimukset (Seurantatutkimukset kunnostuskohteiden laadun ylläpitämiseksi) Ari Huusko Jokien perkaus ja kunnostus Suomessa Luonnontilainen joki --- Intensiivijakso 1930-1960
Kunnostusten seuranta ja seurantatutkimukset (Seurantatutkimukset kunnostuskohteiden laadun ylläpitämiseksi) Ari Huusko Jokien perkaus ja kunnostus Suomessa Luonnontilainen joki --- Intensiivijakso 1930-1960
Espoon kaupungin opetussuunnitelmalinjaukset. Nöykkiön koulu Opetussuunnitelma Biologia. VUOSILUOKAT 7 9 7.lk
 Nöykkiön koulu Opetussuunnitelma Biologia 9.10 a Biologia Oppiaineen opetussuunnitelmaan on merkitty oppiaineen opiskelun yhteydessä toteutuva aihekokonaisuuksien ( = AK) käsittely seuraavin lyhentein:
Nöykkiön koulu Opetussuunnitelma Biologia 9.10 a Biologia Oppiaineen opetussuunnitelmaan on merkitty oppiaineen opiskelun yhteydessä toteutuva aihekokonaisuuksien ( = AK) käsittely seuraavin lyhentein:
Kiintoaineen ja humuksen mallintaminen. Markus Huttunen ja Vanamo Seppänen 11/11/2013
 Kiintoaineen ja humuksen Nitrogen loading from forested catchments mallintaminen Markus Huttunen ja Vanamo Seppänen 11/11/213 Marie Korppoo VEMALA catchment meeting, 25/9/212 21.11.213 VEMALA vedenlaatumalli
Kiintoaineen ja humuksen Nitrogen loading from forested catchments mallintaminen Markus Huttunen ja Vanamo Seppänen 11/11/213 Marie Korppoo VEMALA catchment meeting, 25/9/212 21.11.213 VEMALA vedenlaatumalli
Metsäbioenergian kestävyyden rajat
 Metsäbioenergian kestävyyden rajat Antti Asikainen, professori, METLA Päättäjien metsäakatemia 36. kurssi 07.05.2014 Majvik Storyline Metsä, vedet ja biomassan intensiivinen korjuu Metsien ja metsäenergian
Metsäbioenergian kestävyyden rajat Antti Asikainen, professori, METLA Päättäjien metsäakatemia 36. kurssi 07.05.2014 Majvik Storyline Metsä, vedet ja biomassan intensiivinen korjuu Metsien ja metsäenergian
ytön n vaikutukset vesistöjen ekologisessa tilassa esimerkkinä Muhosjoki
 Näkyvätkö maankäyt ytön n vaikutukset vesistöjen ekologisessa tilassa esimerkkinä Muhosjoki Kaisa Heikkinen 1, Jaana Rintala 1, Satu Maaria Karjalainen 1, Minna Kuoppala 2 & Seppo Hellsten 2 1 Pohjois
Näkyvätkö maankäyt ytön n vaikutukset vesistöjen ekologisessa tilassa esimerkkinä Muhosjoki Kaisa Heikkinen 1, Jaana Rintala 1, Satu Maaria Karjalainen 1, Minna Kuoppala 2 & Seppo Hellsten 2 1 Pohjois
Kohti riistarikkaita reunoja - vaihettumisvyöhykkeiden hoito
 Kohti riistarikkaita reunoja - vaihettumisvyöhykkeiden hoito Piirroakuvat: Jari Kostet ja Tom Björklund. Valokuvat: Sami Tossavainen Vaihettumisvyöhykkeet Tarkoitetaan kahden erilaisen ekosysteemin reuna-
Kohti riistarikkaita reunoja - vaihettumisvyöhykkeiden hoito Piirroakuvat: Jari Kostet ja Tom Björklund. Valokuvat: Sami Tossavainen Vaihettumisvyöhykkeet Tarkoitetaan kahden erilaisen ekosysteemin reuna-