Veren hyytyminen, K-vitamiiniepoksireduktaasi ja varfariini
|
|
|
- Eeva-Liisa Hyttinen
- 7 vuotta sitten
- Katselukertoja:
Transkriptio

1 Kandidaatintutkielma Veren hyytyminen, K-vitamiiniepoksireduktaasi ja varfariini Margareta Kurkela Oulun yliopisto Biokemian ja molekyylilääketieteen tiedekunta 2017
2 Sisällysluettelo Käytetyt lyhenteet Sisällys I KIRJALLISUUSTUTKIELMA 1. Johdanto 4 2. Veren hyytyminen Ulkoinen järjestelmä Sisäinen järjestelmä Hyytymisjärjestelmä tekijästä Xa eteenpäin Gla-karboksyloidut hyytymistekijät Hyytymisjärjestelmän säätely ja inhibointi K-vitamiini K-vitamiinisykli Ihmisen K-vitamiiniepoksireduktaasi hvkor hvkor:n rakenne 15 3TM-malli hvkor:n rakenteelle 16 4TM-malli hvkor:n rakenteelle hvkor:n reaktiomekanismi hvkor:n pelkistäjä Gammakarboksylaasi GGCX GGCX:n rakenne GGCX:n reaktiomekanismi Vasta-aine-entsyymi 20 4.Varfariini Varfariinin sitoutuminen hvkor:aan TM-mallin mukainen sitoutuminen 1-2-TM-lenkkiin TM-mallin mukainen sitoutuminen varfariinia sitovaan taskuun Varfariini-inhibition mekanismi Varfariinin tutkimisen ongelmat In vitro-olosuhteissa Kirjallisuusviitteet 25 II LIITE TIEDETTÄ POPULARISOIVA ESITYS 27 2
3 Käytetyt lyhenteet ADP adenosiinidifosfaatti (adenosine diphosphate) ala alaniini Arg arginiini Asn asparagiini dbsa tioli-disulfidin vaihtajaproteiini ER endoplasmakalvosto (endoplasmic reticulum) gla gammakarboksyloitu glutamaatti GGCX gammakarboksylaasi Gly glysiini his histidiini HMW suurimolekyylipainoinen (high molecular weight) hvkor ihmisen K-vitamiiniepoksireduktaasi Ile isoleusiini K kinoni K1 fyllokinini K2 menakinoni KH2 K-vitamiinihydrokinoni KO K-vitamiiniepoksidi LACI lipoproteiiniin assosioituva hyytymisenestotekijä Leu leusiini lys lysiini MK menakinoni PDI proteiinidisulfidi-isomeraasi phe fenyylialaniini PK fyllokinoni PL fosfolipidi (phospholipid) PTA plasmatromboplastiinikorrelaatti (plasma thromboplastin antecedent) PTC plasmatromboplastiinikomponentti (plasma thromboplastin component) Ser seriini SPCA seerumiprotrombiinikonversiokiihdyttäjä (serum prothrombin conversion accelerator) synvkor synechococcus-bakteerin K-vitamiiniepoksireduktaasi TFPI kudostekijäreitti-inhibiittori (tissue factor pathway inhibitor) TM kalvonläpäisevä (transmebrane) Trp tryptofaani TRX tioredoksiini Tyr tyrosiini VKDP K-vitamiinista rippuvainen proteiini (vitamin K depentent protein) VKOR K-vitamiiniepoksireduktaasi 3
4 1. Johdanto Veren hyytyminen on pitkä reaktioiden sarja, joka lopulta johtaa trombiinin katalysoimana fibriinin muodostumiseen fibrinogeenista, jolloin syntyy verihyytymä, trombi. Aiheettoman trombin syntyminen verisuonen sisään tukkii verisuonia ja paikallisesti haittaa verenkiertoa. Vakavampia seurauksia voi aiheuttaa trombista irtoava osa, joka kulkeutuu verenkierron mukana muualle elimistöön. Trombi voi tukkia sydämen sepelvaltimoita ja saada aikaan sydäninfarktin. Keuhkoihin kulkeutuessaan se aiheuttaa keuhkoembolian. Aivojen verisuonissa syntynyt, tai sinne kulkeutunut trombi voi aikaansaada aivoinfarktin. Antikoagulantteja käytetään lääkkeenä veren liialliseen hyytymiseen. Yleisimmät näistä lääkkeistä ovat varfariini ja hepariini. (Koulu & Tuomisto 2000) Kumariinit löydettiin sattumalta Yhdysvalloissa 1920-luvulla samoihin aikoihin kuin K- vitamiinit. Karjaeläimet söivät pilaantunutta rehua, joka sisälsi paljon dikumarolia, Karl Link tunnisti tämän aineen, ja havaittiin, että vastalääkkeeksi dikumarolin aiheuttamaan hallitsemattomaan verenvuotoon tehosi suuri annos K-vitamiinia. Dikumarolia tutkittiin laboratorioissa ja sen analogeja kehiteltiin useita kymmeniä. Yksi näistä analogeista oli varfariini. Varfariini on ainoa nykyään antikoagulanttina käytetty kumariinijohdannainen. Varfariini ei kuitenkaan suoraan päätynyt lääkeaineeksi, vaan ensin sitä käytettiin rotanmyrkkynä. Varfariinia käytettiin rotanmyrkyissä niin paljon, että se on aiheuttanut massiivisen evoluutiopaineen rottien VKOR-geenille. Vuonna 2009 löydettiin 29 erilaista rotan VKOR:n mutaatiota, jotka aiheuttavat varfariiniresistenssin. Kesti pitkään, ennen kuin varfariinia alettiin käyttää ihmisten hyytymishäiriöiden hoidossa, sillä rotanmyrkyn tärkeintä vaikuttavaa ainetta ei ollut helppo tuoda markkinoille lääkkeenä ihmisille. Nykypäivänä varfariini kuitenkin on yleisin antikoagulantti. Pitkän aikavälin hoidossa se on yleisempi kuin hepariini. Varfariinia alettiin käyttää lääkkeenä 1954, noin 30 vuotta sen löytämisen jälkeen (Van Horn 2013). Nykyään joka sadas Yhdysvaltain ja Iso-Britannian kansalainen käyttää varfariinia säännöllisesti veren hyytymiseen liittyvien sairauksien, kuten keuhkoembolian ja laskimotukoksen sekä sydänkohtauksen ehkäisyyn. Varfariinin yleisyydestä huolimatta sen toimintamekanismi ja kohde-entsyymin rakenne ja toiminta ovat huonosti tunnetut. Parempi ymmärrys vaikutuksesta on tärkeää, sillä varfariinin turva-alue on erittäin kapea. Varfariiniherkkyys myös vaihtelee henkilökohtaisesti, joten yliannostuksen riski on huomattava. (Shen et al. 2017) 4
5 Varfariini on otettu lääkkeenä käyttöön yli 60 vuotta sitten, mutta vasta nyt on alettu ymmärtää sen sitoutumista VKOR:iin ja inhibition mekanismia tarkemmin. Tuloksien avulla voidaan alkaa suunnitella tulevaisuuden turvallisempia varfariinianalogeja antikoagulanteiksi. Lisäksi yliannostuksen riski pienenee mekanismin selvitessä. (Shen et al. 2017) Tutkielmassa käydään laajasti veren hyytymisjärjestelmä läpi ja perehdytään K- vitamiinisykliin kiinnittäen erityistä huomiota ihmisen K-vitamiiniepoksidireduktaasin toimintamekanismiin. Lopuksi käydään läpi varfariinin inhiboiva vaikutusmekanismi K- vitamiiniepoksireduktaasiin. Tällöin saadaan kattava kuva siitä, missä vaiheessa hyytymisjärjestelmää varfariinin inhibitio vaikuttaa. 2. Veren hyytyminen Hemostasia tarkoittaa verenvuodon tyrehdyttämistä. Verisuonen vaurioituessa sen sisäpinnan endoteelisoluja rikkoontuu, mikä aktivoi verihyytymän eli trombin muodostamisen. Vaurion syntyessä verisuoneen vaurioituneen alueen verenkierto minimoituu verisuonen supistuessa. Samalla aktivoituu veren hyytymisjärjestelmä, joka voidaan jakaa kolmeen vaiheeseen: i) Verihiutaleiden, eli trombosyyttien aggregoituminen vauriokohtaan. Vaurion seurauksena paljastuu ekstrasellulaarisia suonenulkoisia kollageenisäikeitä, joihin aktivoituneet trombosyytit sitoutuvat ja alkavat erittää muita trombosyyttejä aktivoivia molekyylejä, ADP:ta ja tromboaksaani A2:ta. Kudosvaurio aktivoi myös hyytymiskaskadin, joka tuottaa trombiinia. Fibrinogeenin aktivoimisen lisäksi trombiini aktivoi myös trombosyyttejä. ii) Fibriiniverkon muodostuminen vauriokohtaan. Verkko muodostaa vauriokohtaan aggregoituneista trombosyyteistä tiiviin hyytymän, trombin. iii)plasmiini liuottaa hyytymän vaurion korjaannuttua. Valtimoihin muodostuvat trombit ovat niin sanottuja valkoisia trombeja, jotka muodostuvat pääosin trombosyyteistä ja fibriinistä. Nopean virtauksen alueilla trombit eivät sisällä erytrosyyttejä eli punasoluja. Punainen trombi muodostuu hitaan verenkierron alueilla laskimoissa, ja se voi muodostua myös ilman verisuonen vauriota. (Granner et al 2006) Trombosyytit aktivoituvat, kun ne joutuvat vaurion myötä kosketuksiin kollageenin kanssa, ja 5
6 kiinnittyvät vauriokohtaan (Koulu & Tuomisto 2000). Trombosyytit muodostavat aloitushyytymän, jonka ympärille erytrosyyteistä ja fibriiniverkosta muodostuva hyytymä kertyy. In vitro-olosuhteissa muodostuva hyytymä on rakenteeltaan samanlainen kuin punainen trombi. Pienimmissä hiussuonissa trombi muodostuu ainoastaan fibriinistä. (Granner et al. 2006) Veren hyytymisjärjestelmän aktivoi verisuonen ulkoisen kudostekijän paljastuminen suonen seinämän vaurioituessa tai negatiivisesti varautuneen pinnan paljastuminen. Hyytymisjärjestelmän aktivointi johtaa reaktioketjuun, jonka lopputuloksena muodostuu aktiivinen trombiini. Se katalysoi fibriinipolymeerien muodostumisen fibrinogeenistä. Hyytymisjärjestelmän tekijöitä on viittä tyyppiä: tsymogeenit, kofaktorit, fibrinogeeni, transglutaminaasi ja säätelevät tekijät (Granner et al. 2006). Tekijät on nimetty yleisen nimensä lisäksi roomalaisilla numeroilla I XIII, mutta numerointi ei noudata järjestystä, sillä tekijät on nimetty löytymisjärjestyksessä, eikä numero VI vastaa mitään hyytymistekijää. Aktiivinen tekijä erotetaan inaktiivisesta lisäämällä a-kirjain tekijän numeron perään, esimerkiksi inaktiivinen fibrinogeeni on tekijä I ja aktiivinen fibriini on tekijä Ia. Hyytymistekijät on esitetty numerojärjestyksessä taulukossa I. 6
7 Taulukko I Veren hyytymistekijät. Hyytymistekijät on esitetty numerojärjestyksessä. Tämä järjestys ei kuitenkaan noudata reaktioiden järjestystä (kuva 1). Tekijöihin I-IV viitataan kirjallisuudessauseimmiten yleisellä nimellään numeron sijasta. Lisäksi taulukossa ovat hyytymistä säätelevät proteiinit C, S ja trombomoduliini. Gammakarboksyloidut tekijät on eritelty taulukossa. Karboksyloidulla proteiinilla on lähellä lähellä aminopäätään useita gammakarboksyloituja glutamaattisivuketjuja. Esimerkiksi trombiinin gla-domeenissa on kymmenen karboksyloitua gutamaattisivuketjua. Gla-domeenin tehtävä on sitoa tekijä negatiiviselle reaktioalustalle Ca 2+ :n kautta. (Granner et al. 2003) Tekijä Yleinen nimi Funktio Järjestelmä γ-karboksylaatio I Fibrinogeeni Verkon rakenneosa Yhdistynyt II Protrombiini Seriiniproteaasi Yhdistynyt Kyllä III Kudostekijä Kofaktori Ulkoinen IV Ca2+ Kofaktori Sisäinen & yhdistynyt V Proakkeleriini Kofaktori Yhdistynyt VII Prokonvertiini Seriinipreoteaasi Ulkoinen Kyllä VIII Antihemofiilinen globuliini IX Antihemofiilinen tekijä B, (Christmas factor) Kofaktori Yhdistynyt Seriiniproteaasi Sisäinen Kyllä X Stuart-Power -tekijä Seriiniproteaasi Yhdistynyt Kyllä XI Plasman tromboplastiinikorrelaatti Seriiniproteaasi Sisäinen XII Hageman tekijä Seriiniproteaasi Sisäinen XIII Fibriiniä stabiloiva tekijä (FSF), fibrinoligaasi transglutaminaasi Yhdistynyt Proteiini C Säätelijä Yhdistynyt Kyllä Proteiini S Säätelijä Yhdistynyt Kyllä Trombomoduliini Säätelijä Yhdistynyt Järjestelmän toiminta perustuu siihen, että edellisen reaktion tuote aktivoituessaan muuttuu proteaasiksi, joka aktivoi seuraavan reaktion tsymogeenin eli proentsyymin, joka aktivoituessaan saa myös proteaasiaktiivisuuden. Proteaasien karboksyylipään aktiivinen keskus on homologinen trypsiinin kanssa, mutta hyytymisjärjestelmän proteaasien spesifistyys on huomattavasti voimakkaampaa kuin trypsiinin. Hyytymisjärjestelmään kuuluu sisäinen ja ulkoinen reitti, joista ulkoinen aktivoituu ensimmäisenä. Reitit yhdistyvät ennen trombiinin 7
8 aktivointia, mutta eivät ole täysin erilliset, sillä ulkoisella reitillä on sisäistä reittiä aktivoiva vaikutus. Hyytymisjärjestelmä kokonaisuudessaan on esitetty kuvassa 1. (Voet et al. 2008) 2.1. Ulkoinen järjestelmä Verisuonen vaurioituessa ensimmäisenä aktivoituu ulkoinen järjestelmä. Ulkoisen järjestelmän tekijöitä ovat kudostekijä, tekijä VII (yleiseltä nimeltään prokonvertiini, seerumiprotrombiinikonversiokiihdyttäjä (SPCA) tai kotromboplastiini) ja tekijä X (yleiseltä nimeltään Sturt-Prower -tekijä). Lisäksi järjestelmän kofaktorina on Ca 2+ (Kuva 1). 8
9 Sisäinen järjestelmä Prekallikreiini HMW kininogeeni Kallikreiini Brandykiniini XII XIIa Ulkoinen järjestelmä XI Ca 2+ XIa Kudostekijä IX Ca 2+ IXa VIII VIIIa VIIa VII V Va X Ca 2+ PL Xa Ca 2+ X Protrombiini Ca 2+ PL Trombiini XIII Fibriini (monomeeri) Fibriini (polymeeri) Fibrinogeeni XIIIa Fibriini (ristisidoksia) Kuva 1: Hyytymisjärjestelmä. Kuvassa on esitetty hyytymisjärjestelmän sisäinen ja ulkoinen hyytymisreitti sekä näiden yhteydet. Siniset nuolet kuvaavat reaktiota ja oranssit nuolet kuvaavat nuolen osoittaman reaktion katalysointia. Kofaktorit Ca 2+ ja PL on merkitty reaktionuolten alle. Hyytymisjärjestelmän aktivoi kudostekijän paljastuminen tai negatiivisen pinnan kanssa kosketuksiin joutunut tekijä. Järjestelmän lopputuotteena syntyy kovalenttisesti ristisidoksia muodostanut fibriinipolymeeriverkko. Järjestelmän edetessä signaali voimistuu, 9
10 eli hyvin pienen määrän järjestelmän alun hyytymistekijän aktivoiminen johtaa moninkertaiseen fibriiniaktivoitumiseen. (Granner et al. 2003) Ulkoisen järjestelmän aktivoitumisen seurauksena tekijä X aktivoituu seriiniproteaasiksi. Ulkoinen järjestelmä aktivoituu verisuonen vaurioituessa, kun ympäröivä kudos joutuu kosketuksiin veren kanssa ja endoteelisolut aktivoituvat tuottamaan pinnalleen kudostekijää, kudostromboplastiinia, joka aktivoi tekijän VII (Koulu & Tuomisto 200, Granner et al. 2003). Kudostekijä on myös muodostuvan tekijän VIIa kofaktori ja tekijät muodostavat kudostekijäkompleksin. Tämän kompleksin muodostuminen on tärkein tapahtuma verenhyytymisjärjestelmän aktivoitumisen ja hyytymänmuodostumisen aloituksen kannalta. Kompleksissa oleva VIIa katalysoi tekijän X Arg-Ile- peptidisidoksen katkeamisen, jolloin tekijä aktivoituu. (Granner et al. 2003) 2.2. Sisäinen järjestelmä Myös sisäisen järjestelmän aktivoitumisen tuloksena tekijästä X muodostuu aktiivinen seriiniproteaasi. Järjestelmän tekijöitä ovat XII (yleiseltä nimeltään Hagemanin tekijä), XI (yleiseltä nimeltään pasmatromboplastiinikorrelaatti (PTA), IX (yleiseltä nimeltään antihemofiilinen tekijä B, Christmas factor tai plasmatromboplastiinikomponentti (PTC), tekijä VII, sekä tekijä X. Lisäksi sisäiseen järjestelmään kuuluu prekallikreiini, suurimolekyylipainoinen kininogeeni (HMW kininogeeni), Kalsiumioni Ca 2+, ja trombosyyttien solukalvon fosfolipidi (PL). Sisäisen järjestelmän aktivoi negatiivisesti varautunut pinta, joka paljastuu tekijöille XII ja XI sekä prekallikreiinille ja HMW kininogeenille (Granner et al. 2003). Tällainen pinta voi olla esimerkiksi kollageeni tai ateroskleroottinen plakki (Koulu & Tuomisto 2000). Sisäinen järjestelmä käynnistyy kallikreiinin tai HMW kininogeenin katalysoimalla tekijän XII proteolyyttisellä aktivoinnilla proteaasiksi XIIa. Se katalysoi prekallikreiinin muutoksen kallikreiiniksi ja tehostaa siten itsensä muodostumista. XIIa katalysoi tekijän XIa muodostumisen XI:sta. Lisäksi se katkaisee HMW kininogeenin, jolloin muodostuu brandykiniiniä, jolla on verisuonia laajentava vaikutus. XIa on proteaasi, jonka kofaktorina toimii Ca 2+ -ioni. XIa katalysoi tekijän IX aktivoinnin IXa:ksi. IXa on seriiniproteaasi, joka katkaisee tekijän X Arg-Ile- peptidisidoksen, jolloin myös muodostuva tekijä Xa on seriiniproteaasi. Reaktio tapahtuu aktivoituneiden trombosyyttien solukalvon ulkopinnalla tenaasikompleksissa, joka koostuu tekijöistä X ja IX, sekä kofaktoreista VIIIa ja Ca
11 VIIIa sitoutuu aktivoidun trombosyytin solukalvon ulkopinnalle fosfolipidin fosfatidyyliseriiniin. Tavallisesti nämä fosfolipidit ovat solukalvon sisäpinnalla, mutta aktivoituneessa trombosyytissä ne siirtyvät solukalvon ulkopinnalle. VIIIa tarjoaa tekijöille X ja IXa reaktioalustan. Lisäksi ulkoisen järjestelmän tekijä VIIa aktivoi kompleksia. (Granner et al. 2003) Sisäisen järjestelmän reaktiot ennen tekijän IXa muodostumista eivät veren hyytymisen edistämisen kannalta vaikuta olevan erityisen merkittäviä, sillä inaktivoivat mutaatiot tekijässä XII, prekallikreiinissa tai HMW kininogeenissa eivät aiheuta hyytymishäiriöitä. Myöskään mutaatio tekijässä XI ei välttämättä aiheuta hyytymishäiriötä. Sisäisen järjestelmän tärkeämpi tehtävä voikin olla fibrolyysin aktivoiminen, kun vaurio on korjattu ja hyytymän liuottaminen on tarpeen. Mainitut tekijät aktivoivat plasminogeenia muuttaen sen plasmiiniksi. Lisäksi kallikreiini aktivoi yksiketjuisen urokinaasin. (Granner et al. 2003) 2.3. Hyytymisjärjetelmä tekijästä Xa eteenpäin Ulkoisen järjestelmän kudostekijäkompleksi ja sisäisen järjestelmän tenaasikomplseksi katalysoivat molemmat saman reaktion, joka on tekijän X Arg-Ile- peptidisidoksen katkaisu. Molemmat reaktiot tapahtuvat solukalvon ulkopinnalla. Tekijän Xa muodostumisesta lähtien sisäinen ja ulkoinen järjestelmä etenevät samaa reaktioreittiä. (Granner et al. 2003) Tekijä Xa katalysoi protrombiinin aktivoitumisen trombiiniksi katkaisemalla kaksi peptidisidosta. Aktiivinen trombiini koostuu kahdesta aminohappoketjusta, joita yhdistää rikkisilta. Reaktio tapahtuu protrombinaasikompleksissa trombosyyttien pinnalla. Kompleksi muodostuu tekijästä Xa ja protrombiinista. Lisäksi kompleksissa kofaktorina on Ca 2+ ja tekijä Va. Samoin, kuin tenaasikompleksissa tekijä VIIIa, protrombinaasikompleksissa tekijä Va tarjoaa reaktiolle alustan kiinnittymällä trombosyyttien pinnalle fosfatidyyliseriinien avulla. Aktiivinen trombiini aktivoi tekijän V muutoksen tekijäksi Va, joka kykenee sitoutumaan trombosyyttien pinnalle mahdollistaen kompleksin muodostumisen, mutta trombiinin konsentraation noustessa tarpeeksi korkeaksi, Va inaktivoituu. Trombiini siis sekä tehostaa, että säätelee itsensä muodostumista. (Granner et al. 2003) Trombiini katalysoi fibrinogeenin muuttumisen fibriiniksi. Inaktiivinen fibriini on pitkänomainen proteiini, joka koostuu kolmesta polypeptidiketjuparista. Fibrinogeenissa aminohappoketjujen päät ovat toisiaan lähellä ja varautuneet voimakkaan negatiivisesti. 11
12 Tämän takia fibrinogeeni onkin erityisen vesiliukoinen. (Granner et al. 2003) Trombiini on seriiniproteaasi, joka katkaisee fibinogeenin polypeptidien aminopäistä spesifisesti neljä Arg- Gly- sidosta, jolloin negatiivisesti varautuneet aspartaatti-ja glutamaattiaminohapot irtoavat leikatun ketjun mukana, minkä seurauksena proteiini menettää vesiliukoisuutensa ja muodostuu fibriinimonomeeri (Koulu & Tuomisto 200, Granner et al. 2003). Trombiinin katalysoiman leikauksen myötä paljastuu monomeerin sitoutumiskohta ja alkaa spontaani fibriinipolymeerin muodostuminen. Polymeeriin takertuu erytrosyyttejä ja trombosyyttejä ja muodostuu löyhästi rakentunut hyytymä, jonka rakennetta ylläpitää fibriinimonomeerien väliset heikot sidokset. (Granner et al. 2003) Trombiini katalysoi myös tekijän XIII, eli transglutaminaasin, aktivointia. Transglutaminaasi on entsyymi, joka katalysoi fibriinien sisältämien glutamiinin ja lysiinin välille muodostuvia peptidisidoksia. Transglutaminaasi liittää fibriinimolekyylit kovalenttisesti toisiinsa, jolloin muodostuu vahva fibriiniverkko ja trombi, eli hyytymä. (Granner et al. 2003) Ulkoisen ja sisäisen järjestelmän välillä on yhteys jo ennen tekijää X, sillä ulkoisen järjestelmän tekijä VIIa kykenee katalysoimaan sisäisen järjestelmän tekijän IX aktivoitumisen. (Granner et al. 2003) 2.4. Gla-karboksyloidut hyytymistekijät Protrombiini, tekijät VII, IX ja X sekä proteiinit C ja S ovat gla-karboksyloituja. Gla-domeeni katkaistaan aktivoinnissa protrombiinista. Gla-domeeni koostuu useasta gammakarboksyloidusta glutamaattiaminohaposta, ja domeenilla on suuri affiniteetti sitoa Ca 2+ -ioneja. Gla-domeenin tehtävä on kiinnittää hyytymistekijöitä reaktioalustalle, kuten negatiivisesti varautuneelle trombosyytin solukalvolle kalsiumionien kautta. Jos hyytymistekijöiden karboksylaatio häiriytyy, menettää myös hyytymisjärjestelmä tehoaan (Granner et al. 2003). Näiden tekijöiden karboksylaation katalysoi gammakarboksylaasi, jonka kofaktorina on pelkistynyt K-vitamiini. Karboksyloidut hyytymistekijät ovatkin K-vitamiinista riippuvasia proteiineja (VKDP). (Van Horn 2013) 2.5. Hyytymisjärjestelmän säätely ja inhibointi Hyytymisjärjestelmä on tarkasti säädelty, jotta liiallinen hyytyminen ja haitallisten trrombien muodostuminen olisi estetty. Hyytymisjärjestelmän jokainen reaktio katalysoi seuraavan 12
13 proteaasin aktivoitumista. Reaktioketjun edetessä sen tuotteet säätelevät jatkuvasti aiempien reaktioiden nopeutta varmistaen, ettei hyytymisjärjestelmä aktivoidu liian paljon. Hyytymisjärjestelmän ylävirran tekijöiden konsentraatio on huomattavasti matalampi kuin lopputuotteiden, trombiinin ja fibriinin, eli hyytymissignaali voimistuu edetessään (Granner et al. 2003). Hyytymän muodostuessa aktivoituu myös hyytymää liuottava fibrolyyttinen järjestelmä. Sen tarkoitus on estää trombin kasvaminen ja hajottaa se vauriokohdan endoteelisolujen parantuessa. Kudokset erittävät parantuessaan plasminogeeniaktivaattoria, joka muuttaa veren plamsinogeenia plasmiiniksi. Plasmiini liuottaa fibriiniä, jolloin trombi ei enää pysy kasassa ja liukenee. (Koulu & Tuomisto 2000, Mathews 2000) Antitrombiini on luonnollinen trombiininestäjä, mutta se inhiboi myös tekijöiden IXa, Xa, XIa, XIIa ja kudostekijäkompleksin aktiivisuutta. Toinen luonnollinen inhibiittori hyytymisjärjestelmälle on kudostekijäreitti-inhibiittori (TFPI), joka sitoutuu aktiiviseen tekijään Xa lähelle sen aktiivista keskusta, ja muodostuva kompleksi inhiboi kudostekijäkompleksia. TFPI:n inhiboiva vaikutus perustuu paitsi Xa:n inaktivoimiseen, myös sen muodostumisen estämiseen. (Granner et al. 2003) Muita luonnollisia antikoagulantteja ovat esimerkiksi prostasykliini, joka estää trombosyyttien aggregaation. Plasman antitrombiini III inaktivoi hyytymistekijöitä IIa, IXa, XIa ja XIIa. Endoteelisolut erittävät hepariinisulfaattia, joka stimuloi antitrombiinia. Aktivoitu proteiini C pilkkoo tekijöitä Va ja VIIa, kun proteiinia S on läsnä. Trombiini, trombomoduliini ja tekijän gammakarboksylaatio aktivoivat proteiini C:n. Myös tämä karboksylaatio on riippuvainen pelkistetystä K-vitamiinista, joten K-vitamiini aktivoi paitsi hyytymistekijöitä, myös niitä estäviä tekijöitä. LACI on lipoproteiiniin assosioituva hyytymisenestotekijä, joka estää hyytymistekijää Xa ja VII-kompleksia muodostumasta. (Koulu & Tuomisto 2000) 3. K-vitamiini Henrik Dam selvitti K-vitamiinin rakenteen vuonna K-vitamiinit ovat yhdisteryhmä, joille yhteistä on voimakas hydrofobisuus. Kaikilla K-vitamiinityypeillä on sama pääryhmä, 2- metyyli-1,4-naptokinoni, mutta 3-hiileen kiinnittyneen sivuketjun rakenne vaihtelee. Kuten muitakaan vitamiineja, ihmisen keho ei itse kykene tuottamaan tarvitsemaansa K-vitamiinia, vaan se on saatava ravinnosta tai muista lähteistä. Pääasiallinen K-vitamiinin lähde on 13
14 kasviperäinen fyllokinoni (PK tai K1), joka kasveissa on elektroninsiirtoketjun kofaktori (Van Horn 3013). PK muokataan kehossa menakinoniksi MK4 (Koulu & Tuomisto 2000, Van Horn 2013). Paksusuolen bakteerien tuottama K-vitamiini on menakinonia (K2 tai MK). (Koulu & Tuomisto 2000). Menakinonin sivuketju on isopreeniketju, jossa on korkeintaan 12 isopreeniyksikköä. Bakteerit eivät kuitenkaan syntetisoi neljää isopreeniyksikköä sisältävää menakinonia MK4. Ravinnosta saatu K-vitamiini metaboloidaan MK4:ksi (Van Horn 2013). Paksusuolen bakteerit syntetisoivat noin puolet kehon tarvitsemasta K-vitamiinimäärästä, mutta fyllokinoni on silti tärkein K-vitamiininlähde, sillä menakinonin imeytyminen suolistosta on tehotonta ja riittämätöntä (Voet et al. 2008). Jos ruokavalio ei sisällä tarpeeksi K-vitamiinin lähteitä, puutosoireita voi ilmetä, vaikka suoliston bakteerit tuottavatkin sitä (Granner et al. 2003). Myös synteettiset menadinoni ja menadioli ovat aktiivisia, sillä ne metaboloidaan elimistössä käyttökelpoiseen muotoon MK4:ksi (Granner et al. 2003, Voet et al. 2008). K-vitamiinin tärkein tehtävä on maksassa tapahtuvan veren hyytymistekijöiden gammakarboksylaation mahdollistaminen. (Koulu & Tuomisto 2000) Nimitys K-vitamiini juontuukin saksan kielen sanasta Koagulation, joka tarkoittaa hyytymistä (Voet et al. 2008). K- vitamiini on gammakarboksylaasin kofaktori. Tämä entsyymi katalysoi hyytymistekijöiden glutamaattien gammakarboksylaation (Koulu & Tuomisto 2000). Glutamaatin gammakarboksylaatio on harvinainen posttranslationaalinen muokkaus, jota esiintyy veren hyytymistekijöiden kalsiumia sitovissa domeeneissa. (Granner et al. 2003) K-vitamiinin puutos johtaa joko vajavaisesti karboksyloituihin tai täysin karboksyloitumattomiin hyytymistekijöihin, joilla siten gla-domeeni toimii heikosti tai on täysin toimimaton. Tällöin hyytymistekijät eivät voi sitoutua negatiivisesti varautuneelle reaktioalustalle ja hyytymisketju pysähtyy (Voet et al. 2008). K-vitamiinin avulla karboksyloidaan myös muita kalsiumia sitovia proteiineja, kuten luuhomeostasiaan vaikuttavia ja oksidatiiviseen sterssiin reagoivia proteiineja, mutta niihin ei tässä tutkielmassa perehdytä. (Van Horn 2013) 3.1. K-vitamiinisykli K-vitamiinisykli koostuu pelkistyneestä hydrokinonista KH2, hapetetusta K- vitamiiniepoksidista KO sekä K-vitamiinikinonista K tai MK4. Ravinnosta saatava fyllokinoni metaboloidaan menakinoniksi MK4, joka pelkistetään aktiiviseksi hydrokinoniksi KH2. Pelkistäjänä on K-vitamiiniepoksireduktaasi VKOR tai vasta-aine-entsyymi (antidotal enzyme). Aktiivinen KH2 on gammakarboksylaasin GGCX kofaktori. GGCX karboksyloi 14
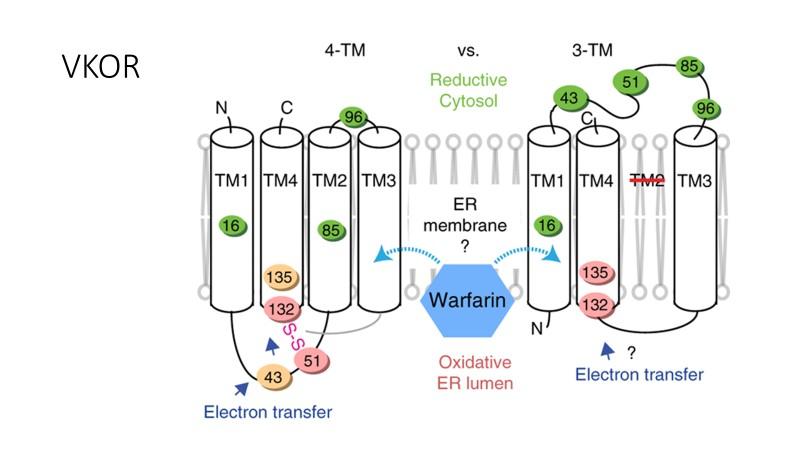
15 K-vitamiinista riippuvaisten proteiinien gla-domeenin gluamaatteja hapettaen samalla KH2:n hydrokinoniksi KO. VKOR pelkistää KO:n MK4:ksi, ja sykli voi edetä uudelle kierrokselle. (Van Horn 2013) Ihmisen K-vitamiiniepoksireduktaasi hvkor hvkor:n ensisijainen tehtävä on katalysoida KO:n pelkistäminen K:ksi, ja toissijainen tehtävä on K:n pelkistäminen aktiiviseksi KH2:ksi. (Tie et al 2011) hvkor:n rakenne Vuonna 2004 Oldenburg ja Stafford selvittivät ihmisen K-vitamiiniepoksireduktaasia (hvkor) koodaava geenin. Geenin koodaama proteiini on pieni. Se on vain 163 aminohapon pituinen (18kDa). Tämän geenin koodaama entsyymi pystyy kuitenkin itsenäisesti pelkistämään KO:ta K:ksi ja K:ta KH2:ta Ksi. Tästä johtuen VKOR ei välttämättä ole osa suurempaa kompleksia (Li et al 2004). hvkor on transmembraaniproteiini, jonka muodostaa kolme tai neljä endoplasmakalvoston ER:n läpäisevää alfakierrettä (kuva 2). Molempia rakenteita tukevia tutkimustuloksia on julkaistu (Goodstadt & Ponting 2004, Tie et al. 2016). Entsyymin aktiivisen keskuksen CxxC:n muodostavat kysteiinit cys132 ja cys135. Kaikkien eliöiden VKOR:ssa on neljä täysin konservoitunutta kysteiiniä. hvkor:ssa nämä ovat cys43, cys51, cys132 ja cys135. Lisäksi hvkor:n ser57 on konservoitunut kaikissa VKOR:ssa seriiniksi tai treoniiniksi. (Van Horn 2013) Substraatit KO ja K ovat voimakkaasti hydrofobisia, joten on loogista, että VKOR:n aktiivinen keskus CxxC, joka on homologinen tiolinpelkistäjäentsyymien kanssa, sijaitsee ER:n sisällä hydrofobisissa oloissa. Molempien mallien mukaan aktiivinen keskus sijaitsee viimeisessä alfakierteessä (Goodstadt & Ponting 2004, Li et al. 2010). Bakteerin synechococcus epoksireduktaasin, synvkor, rakenne on määritetty 4TMrakenteeksi, ja sen oletetaan muistuttavan ihmisen VKOR:n rakennetta paljolti, teoriat hvkor:n rakenteesta perustuvat paljolti sovellettuina synvkor:n rakenteeseen. Proteiinit kuitenkin eroavat toisistaan melko paljon, joten tämän perusteella ei hvkor:n rakennetta voi varmuudella määrittää (van Horn 2013). 15

16 Kuva 2. Vaihtoehtoiset mallit hvkor:n rakenteelle. Vihreät kysteiinit ovat pelkistyneinä, oranssit osittain pelkistyneinä ja vaaleanpunaiset lähinnä hapettuneessa muodossa. Nuolet esittävät mahdollista reittiä elektronin kuljetuksesta pelkistäjältä substraatille. Vasemmalla on malli 4TM rakenteesta, jossa kysteiinien 51 ja 132 välille muodostuu rikkisilta. Oikealla malli on 3TM rakenteesta, jossa pelkistävä rikkisilta muodostuu mahdollisesti TM3 alfakierteestä. Varfariinin sitoutumista entsyymiin käsitellään tutkielmassa myöhemmin (Lähde Shen et al 2017, Nature. Kuvan käyttöön on julkaisijan lupa.) 3TM-malli hvkor:n rakenteelle 3TM Teorian mukaan entsyymi koostuu kolmesta ER:n läpäisevästä alfakierteestä (Kuva 2) ja niitä yhdistävistä lenkeistä. Entsyymin C-termilaaliosa jää sytosoliin ja N-terminaali ER:n lumeniin. Mallin mukaan aktiivinen keskus, joka sisältää kysteiinit 132 ja 135, sijaitsee ER:n sisällä lähellä lumenia. 1-2-TM -lenkki, joka sisältää konservoituneet aminohapot cys43, cys51 ja ser57, on teorian mukaan sytosolissa. Kysteiinien cys43 ja cys51 on esitetty osallistuvan elektronin siirtämiseen K-vitamiinille. 3TM-rakenne ei tue tätä reaktiomekanismia, sillä se sijoittaa nämä kysteiinit sytosoliin ja aktiivisen keskuksen ER:n lumenin puolelle, eli akenteet sijaitsvat eripuolella kalvoa.. Jos 3TM malli on oikea, on mahdollista, että gammakarboksylaasi 16
17 GGCX muodostaa kompleksin hvkor:n kanssa. (Li et al 2004) 3TM-malli ei kykene tyydyttävästi selittämään varfariini-inhibition mekanismia, jota käsitellään tutkielmassa myöhemmin. Todetaan kuitenkin, että mutaatio cys51:ssa aiheuttaa varfariiniresistenssin. Tätä kysteiiniä on esitetty varfariinin sitoutumiskohdaksi (Tie et al 2016). Varfariini on kuitenkin voimakkaasti hydrofobinen ja kyseinen kysteiini sijaitsee 3TM-mallin mukaan hydrofiilisissä oloissa sytoplasmassa. Positiivisesti varautuneet aminohapot perinteisesti proteiinirakenteissa sijaitsevat kalvojen sisäpuolella, mikä toteutuu 3TM-mallissa hvkor:lle (Van Horn 2013). 4TM-malli hvkor:n rakenteelle 4TM-mallin mukaan entsyymin rakenteessa on 4 ER:n läpäisevää alfakierrettä (kuva 2) sekä niitä yhdistävät lenkit. Tämän teorian mukaan myös N-terminaali on sytoplasmassa, mutta aktiivinen kohta sijaitsee ER:n sisällä hydrofobisissa olosuhteissa lähellä lumenin puolta, kuten 3TM-mallissa (Goodstadt & Ponting 2004). Tärkein ero 3TM-malliin on 1-2- TM-lenkin sijainti. 4TM-mallin mukaan kyseinen lenkki on ER:n lumenissa. 4TM-teoriaa tukee kaikkien konservoituneiden aminohappojen sijainti samalla puolella ER:a. hvkor:lle on esitetty reaktiomekanismia, jossa kysteiinit cys43 ja cys51 osallistuvat elektronin siirtoon pelkistäjältä K-vitamiinille (Goodstadt & Ponting 4004, Van Horn 2013). Tämänkaltainen reaktiomekanismi on mahdollinen vain, jos sekä 1-2- TM-lenkki ja aktiivinen keskus sijaitsevat samalla puolella ER:a. (Schulman et al 2010). Goostadt ja Ponting havaitsivat, että cys51 ja cys132 muodostavat rikkisillan. 3TM-malli ei tue kyseisen rikkisillan muodostumista (Goodstadt & Ponting 2004) hvkor:n reaktiomekanismi Tyydyttävää selitystä 3TM-mallin mukaiselle hvkor:n reaktiomekanismille ei ole esitetty. On spekuloitu, että entsyymi muodostaa kompleksin GGCX:n kanssa, mutta koska tästä ei ole esitetty tarkempaa todennäköistä teoriaa reaktion mekanismille, keskitytään tutkielmassa 4TM:n malliin reaktiomekanismille. Aktiivinen entsyymi muodostaa rikkisillan kysteiinien cys51 ja cys132 välille. Nämä kysteiinit ovat solussa 90 %:sti hapettuneita, eli lähes kaikki ovat muodostaneet rikkisillan. Kysteiinit cys43 ja cys135 ovat 45 %:sti hapettuneita. Muut kysteiinit cys16, cys85 ja cys96 ovat pelkistyneitä, mikä selittyy niiden sijainnilla solukalvon läpäisevän alfakierteen keskellä tai 17
18 sytosolin puolella (Goodstadt & Ponting 2004). Pelkistäjä luovuttaa elektronin cys51:lle, joka luovuttaa rikkisillan elektronit cys132:lle. Trp122 deprotonoi substraatin (KO tai K) nukleofiilisella hyökkäyksellä, ja muodostuu karbanioni. Pelkistynyt cys132 muodostaa substraatin 2-hiilen kanssa kovalenttisen sidoksen. Cys132 luovuttaa sidoksen elektronit substraatille, joka pelkistyy. Samalla entsyymi muodostaa rikkisidoksen uudestaan. 3TMmallin mukaiselle rakenteelle on esitetty, että rikkisidos muodostuu GGCX:n jonkin kysteiinin ja hvkor:n cys132:n välille. (Van Horn 2013) Mutaatio kysteiineissä cys132 tai 135 inaktivoi hvkor:n täysin. Myös cys43 ja cys51 mutaatio hidastaa pelkistysreaktiota, mutta ei yhtä voimakkaasti. Aktiivisen keskuksen kysteiinit ovat siis korvaamattomia ja konservoituneet kysteiinit cys43 ja cys51 osallistuvat pelkistymisen katalysointiin, mutta eivät ole välttämättömiä (Jin et al. 2007). Mahdollisesti rikkisillan voi muodostaa myös jokin muu entsyymin kysteiini, vaikka cys51 on ensisijainen rikkisillan muodostaja, sillä cys51 ei vaikuta olevan reaktiolle välttämätön. (Li et al 2017) hvkor:n pelkistäjä Myöskään elektronin luovuttavaa pelkistäjää ei ole selvitetty. Todennäköisin pelkistäjä on ER:n kalvoproteiini tioredoksiini (TRX) tai jokin muu tioli. Pelkistäjäksi on myös ehdotettu TRX:n kaltaista oksidoreduktaasia, proteiinidisulfidi-isomeraasia (PDI) ja tioli-disulfidivaihtajaproteiinia (dsba), mutta vakuuttavia tuloksia ei ole saatu (Goodstadt & Ponting 2004, Van Horn 2013). Kaikki mahdolliset pelkistäjät kykenevät muodostamaan rikkisillan oman aktiivisen CxxC-keskuksensa kautta hvkor:n kanssa, mitä kautta elektroninluovutus tapahtuisi. TRX muodostaa hanakaimmin rikkisidoksia, mikä lisää todennäköisyyttä, että se on pelkistäjä (Shen et al 2017) Gammakarboksylaasi GGCX Gammakarboksylaasi on ER:lla sijaitseva transmembraaninen proteiini, jonka aktiivinen keskus sijaitsee ER:n lumenissa. Se katalysoi VKD-proteiinien glutamaattien gammakarboksylaation. (Rishavy et al 2011) Gammakarboksylaasi on lisäksi epoksidaasi, sillä glutamaatin sivuketjun karboksylaation lisäksi se sivutuotteena hapettaa KH2:ta muodostaen KO:ta (Mathews et al. 2000). Gammakarboksylaasin toinen nimi onkin K-vitamiiniepoksidaasi. K-vitamiinisyklistä 18
19 puhuttaessa käytetään yleisesti tätä nimeä, mutta kun keskitytään VKDP:n karboksylaatioon, käytetään nimitystä gammakarboksylaasi (Granner et al. 2003). Gammakarboksylaasi käyttää kofaktorinaan KH2:a. GGCX katalysoi glutamaatin gammakarboksylaation, jossa KH2 deprotonoi glutamaatin C-4 -hiilen. Sivutuotteena syntyy hapettunutta KO:ta (Mathews et al. 2000). Tämän seurauksena CO2 pystyy spontaanisti, ilman entsyymikatalyysiä hyökkäämään syntyneen glutamaattikarbanioni sivuketjun C-4 hiileen, jolloin syntyy karboksyloitu glutamaatti (gla) (Mathews et al. 2000, Granner et al. 2003). Gammakarboksylaasin katalysoima VKDP:n karboksylaatio on riippuvainen KH2:ta. Jos karboksylaatio ei toimi, syntyy vain osittain karboksyloituneita tai täysin karboksyloitumattomia proteiineja, joilla on vajavainen kyky tai ei kykyä lainkaan sitoa kalsiumioneja ja kiinnittyä veren hyytymistekijöiden tapauksessa reaktiopinnalle. Karboksylaation inaktivointi siis estää hyytymisketjun toimintaa. karboksyloidut hyytymistekijät ovat trombiini sekä tekijät VII, IX ja X GGCX:n rakenne Karboksylaasin rakennetta ei ole saatu kunnolla ratkaistua, mutta tiedetään, että se on ER:lla sijaitseva kalvoproteiini, jonka aktiivinen keskus on lumenin puolella. Lys218 on katalyyttinen emäs, joka deprotonoi KH2:n ja aloittaa reaktion. Täysin konservoitunut aminohappo his160 kytkee epoksidaation substraatin aminohappojen karboksylaatioon. Mutaatio, joka muuttaa tämäm histidiinin alaniiniksi, katalysoi kyllä KH2:n epoksidaation KO:ksi, mutta mutatoitunut entsyymi ei pysty karboksyloimaan glutamaattia. His160:n tehtävä on helpottaa karbanioidun glutamaatin reaktiota hiilidioksidin kanssa. Kun His160 korvattiin alaniinilla (ala), karbanioitu glu-sivuketju protonoitui todennäköisemmin kuin reagoi hiilidioksidin kanssa. (Rishavy et al. 2008) GGCX:n reaktiomekanismi Karboksylaatioon GGCX saa elektronit K-vitamiinin epoksidaatiosta, jossa KH2 hapettuu KO:ksi. Mekanismia ei tarkkaan tunneta, mutta tutkimukset viittaavat siihen, että karboksylaatio tapahtuu niin sanotulla emäksen vahvistamismenetelmällä, jossa GGCX:n aktiivinen keskus his 160 katalysoi KH2:n deprotonaation KH - :ksi. GGCX hapettaa KH - :n, jolloin muodostuu reaktiivinen välituote K -. K - on korkeaenerginen, ja se reagoi hanakasti 19
20 substraatin glutamaattisivuketjun kanssa. Se vastaanottaa glutamaatin gammapaikasta vedyn hapettuen itse KO:ksi. Reaktiossa muodostuu glutamaattikarbanioni, joka reagoi hiilidioksidin kanssa muodostaen karboksyloidun glutamaatin. Yksi KH2 pystyy karboksyloimaan yhden glutamaatin. VKD-proteiinit sisältävät useita gla-sivuketjuja, ja yhtä monta KH2:a tulee pelkistää aktiivisen proteiinin muodostamiseksi (van Horn 2013) Vasta-aine-entsyymi Mutaatio kysteiineissä cys132 tai cys135 inaktivoi hvkor:n täysin. In vivo on havaittu, että tällaisen mutaation vaikutuksesta KO:n pelkistämistä K:ksi ei tapahdu. Kuitenkin on huomattu, että mutaatio ei täysin estä in vivo K:n pelkistämistä aktiiviseksi KH2ksi, vaan jonkin verran aktiivista hydrokinonia muodostuu mutaatiosta huolimatta. In vitro kokeet ovat kuitenkin osoittaneet, että mutaatio aktiivisen kohdan kysteiineissä inaktivoi hvkor:n täysin, eikä se katalysoi kumpaakaan reaktiota. Tästä on päätelty, että myös jokin toinen entsyymi katalysoi ravinnosta saatavan kinonin pelkistämisen hydrokinoniksi tarpeeksi suuren kinonipitoisuuden myötä. Entsyymiä kutsutaan vasta-aine-entsyymiksi, sillä se mahdollistaa varfariiniyliannostuksen hoidon suurella K-vitamiiniannoksella. Kääntöpuolena on myös sen vaikutus varfariiniannosteluun, sillä ravinnosta saatava K-vitamiini voidaan osittain metaboloida aktiiviseksi hydrokinoniksi. Tästä seuraa se, että varfariinia ottavien henkilöiden on suotavaa yrittää syödä tasaisen paljon K-vitamiinia, jotta ravinnosta saatava K-vitamiini ei vaihtelisi paljon. Vasta-aine-entsyymistä ei tiedetä juurikaan mitään, mikä vaikeuttaa varfariiniannostelua, sillä yksittäisen potilaan vasta-aine-entsyymin aktiivisuutta on vaikea arvioida. (Tie et al. 2011). Tämä voi johtaa varfariinin yliannostukseen, jonka seurauksena on hallitsematon sisäinen ja ulkoinen verenvuoto. (Granner et al. 2003) 4. Varfariini Varfariini on kumariinijohdannainen 4-hydroksikumariini, joka on ainoa yhä käytössä oleva antikoagulanttina käytettävä kumariini (Franchini et al. 2016). Varfariini estää glutamaattisivuketjujen karboksylaation. Näitä gla-sivuketjuja on esimerkiksi veren kalsiumia sitovien hyytymistekijöiden (trombiini sekä tekijät VII, IX ja X) sekä veren hyytymistä säätelevien tekijöiden (proteiinit C ja S) gla-domeeneissa. Varfariini ei kuitenkaan suoraan inhiboi näiden gla-domeenien muodostamista, vaan estää gammakarboksylaasin kofaktorin, 20
21 KH2:n, muodostumisen pelkistyneestä KO:sta ja K:sta, inaktivoiden VKOR:n. Tämän epäsuoran inhibition takia varfariinin vaikutus ilmenee vasta, kun sekä valmiit hyytymistekijät että aktiivinen KH2 on kulutettu loppuun. Tähän kuluu yhdestä kahteen vuorokautta, ja se saavuttaa maksimitehonsa viikon kuluessa (Granner et al. 2003) Varfariinin sitoutuminen VKOR:iin Varfariinin sitoutumiskohdaksi on esitetty kahta eri kohtaa hvkor:ssa: TM1-2-lenkin konservoituneet aimohapot cys43 ja cys51 voivat muodostaa varfariinin sitoutumiskohdan. (Tie et al 2016). Toisen teorian mukaan varfariini sitoutuu TM4:ssä aminohappoihin thr138, tyr139 sekä ala140. (Rost et al. 2005). Kumpikaan sitoutumisteorioista ei kykene selittämään ongelmitta inhibitiomekanismia TM-mallin mukainen sitoutuminen 1-2-TM-lenkkiin hvkor:n 3TM rakenteen mukaan varfariini sitoutuu 1-2TM lenkin konservoituneisiin kysteiineihin sytosolissa. Mutaatio aminohapossa cys51 aiheuttaa erittäin voimakkaan varfariiniresistenssin. Lenkki kuitenkin sijaitsee sytosolissa, ja varfariini on voimakkaasti hydrofobinen yhdiste, joten luonnollinen sitoutumiskohta sille olisivat entsyymin ER:n sisäiset rakenteet. Tämä teoria ei myöskään kykene selittämään inhibitiomekansimia. Kysteiinit cys43 ja cys51 eivät ole kovin lähekkäin, joten varfariini ei kykene sitoutumaan molempiin yhtä aikaa. Tiedetään, että vaihtoehtoinen rikkisilta cys43-cys132 voi muodostua, jos cys51 ei rikkisiltaa muodosta, joten varfariinin cys51-cys132 rikkisiltaa estävä vaikutus on epätodennäköinen. Rikkisillan cys43-cys132 muodostuessa varfariini ei kykene inhiboimaan entsyymiä (Rishavy et al. 2011, Tie et al. 2016) TM-mallin mukainen sitoutuminen varfariinia sitovaan taskuun TM4:ssä aminohapot thr138, tyr139 ja ala140 sitovat varfariinin (Rost et al 2005). Tyr139 sijaitsee yhden täyden alfakierroksen verran aktiivisen keskuksen cys132 yläpuolella. Tämän kysteiinin on myös todettu muodostavan rikkisillan cys51:n kanssa. Tämä rikkisidos on erittäin tärkeä varfariinin sitoutumiselle, sillä häiriö sen muodostumisessa johtaa varfariiniresistenssiin (Goodstadt & Ponting 2004). Rikkisidos on varfariinin sitoutumiselle paljon tärkeämpi kuin entsyymin aktiivisuudelle, sillä vaikka cys51 mutatoituisi, kykenee vaihtoehtoinen rikkisidos muodostumaan. Varfariinin sitoutumisessa rikkisidoksen merkitys on varfariinin 21
22 sitoutumistaskun muodostuminen, sillä kyseinen rikkisidos muuttaa entsyymin konformaatiota siten, että varfariinille otollinen hydrofobinen sitoutumiskohta muodostuu. Taskua ympäröivät TM1:n aminohapot gly2, leu22, ser23, leu24, tyr25 ja ala26 sekä useat muut alaniinit. TM4 ja TM1 ovat esitetyn rakenteen mukaan hyvin lähekkäin. TM4-mallin sitoutuminen esitetty kuvassa 3. Tutkimalla varfariiniresistenttejä mutaatiota, varfariinin sitoutumiskohdaksi on esitetty varfariinia sitovaa taskua hvkor:n aktiivisen keskuksen lähellä. Kysteiinien cys43 ja cys132 välille muodostuva rikkisidos muodostaa taskun. Pistemutaatio näissä aminohapoissa aiheuttaa varfariiniresistenssiä. 1-2-TM lenkin aminohappojen phe55 ja arg80 uskotaan sitovan varfariinin sivuketjua (Shen et al. 2017) Varfariinin on esitetty sitoutuvan pelkistyneeseen hvkor:aan,, mutta rikkisilta cys51-cys132 on muodostunut hapettuneessa hvkor:ssa, mikä viittaisi mahdollisuuteen, että varfariini sitoutuukin hvkor:n hapettuneeseen muotoon. (Shen et al 2017) 22

23 Kuva 3. Varfariinin sitoutuminen hvkor:iin 4TM mallin mukaan. Kuviin on merkitty varfariinia sitovaa taskua ympäröivät aminohapot, joiden mutaatiosta seuraa vaihtelevan asteista varfariiniresistenssiä. Rikkisilta cys51-cys132 esitetään kuvissa kirkkaan vihreänä. Kuvassa a) esitetään varfariinin sitoutumistasku. Rikkisillan cys51-cys132 ansiosta TM-1-2- lenkin alfakierre asettuu taskun päälle muodostaen hydrofobisen sitoutumispaikan. Kuvissa b) ja c) esitetään tarkemmin taskua ympäröivät aminohapot ja varfariinia sitova tyr139 TM4:ssä ja sivuketjua sitovat phe55 ja asn80 TM-1-2 -lenkissä. (Lähde Shen et al Nature. Kuvan käyttöön on lupa julkaisijalta.) 23
24 4.2. Varfariini-inhibition mekanismi Varfariini sitoutuu tyr139:aan lähelle aktiivista kohtaa, jolloin sen inhibitiovaikutus voi perustua kahteen seikkaan: elektroninsiirron häiritsemiseen pelkistäjältä substraatille (KO tai K), tai substraatin sitoutumisen estämiseen, jopa sen syrjäyttämiseen. (Shen et al 2017) Varfariini todennäköisesti estää elektroninluovutuksen kysteiinille cys51 tai kysteiiniltä cys51 kysteiinille cys132, jolloin KO:n ja K:n pelkistäminen estyy, koska elektronia ei luovuteta. (Shen et al 2017) Varfariinia sitova tyr139 on niin lähellä aktiivista substraattia sitovaa keskusta, että varfariini ja substraatti eivät mahdu yhtä aikaa sitoutumaan proteiiniin. Toisaalta varfariini-inhibition on kuitenkin todettu olevan nonkompetetiivista, eli substraatin sitoutumisella ei pitäisi olla vaikutusta varfariinin sitoutumiseen, mikä on ristiriidassa tämän sitoutumiskohtien läheisyyden kanssa (Shen et al. 2017). Sitoutumiskohdan selvittäminen auttaa ymmärtämään inhibitiomekanismin ja auttaa lääkkeen kliinisessä käytössä annostuksen määrittämistä. Varfariininyliannostus aiheuttaa voimakasta hallitsematonta verenvuotoa ja haavan syntyessä kyvyttömyyttä hyytymistekijöiden aktivoitumiseen. (Shen et al. 2017) Varfariinin tutkimisen ongelmat In vitro-olosuhteissa In vitro tutkimuksissa hvkor on enemmän pelkistyneessä muodossa kuin solussa, joten siinä ei ole yhtä paljon rikkisidoksia. Lisäksi tutkimuksissa käytetty pelkistäjä DTT häiritsee cys51:n ja cys132:n muodostamaa rikkisiltaa, mikä johtaa siihen, että varfariinin teho heikkenee rikkisillan menetyksen takia. In vitro 50 % inhibitiokonsentraatio on 100-kertainen verrattuna In vivoon. Silti varfariinia on tutkittu pääasiassa In vitro olosuhteissa. (Shen et al 2017) Teorian, jonka mukaan varfariini sitoutuu 1-2 -TM-lenkin kysteiineihin, perusteella ei kyetä selittämään inhibitiomekanismia. On todettu, että varfariini sitoutuu pelkistyneeseen hvkor:aan. Tämä tukee sitoutumista esimerkiksi kysteiiniin cys51, sillä rikkisidoksen muodostuminen estyy, mutta hydrofiilinen olosuhde ei ole otollinen varfariinin sitomiseen. (Tie et al 2016) Myös aktiivisen keskuksen viereen sitoutuminen on ristiriidassa inhibition aikaisempiin ominaisuuksiin. Inhibiton on todettu olevan nonkompetetiivista, mutta varfariinin sitoutuessa 24
25 tyr139:aan, ei substraatti mahdu sitoutumaan, eivätkä ne siten kykene sitoutumaan itsenäisesti. Lisäksi varfariini sitoutuu pelkistyneeseen hvkor:aan, jossa cys51-cys132 rikkisilta on auennut. Tämän sitoutumismekanismin perustana on juuri kyseisen rikkisillan muodostama tasku. Sekä entsyymin rakenne ja reaktiomekanismi, että varfariinin toiminta tunnetaan liian huonosti, että tyydyttävä teoria inhibitiomekanismista voitaisiin esittää. Tutkimus on erityisen tärkeää, sillä paremmalla varfariinin toiminnan ymmärryksellä varfariinin käytöstä tulisi turvallisempaa (Shen et al 2017) 5. Kirjallisuusviitteet Becker RC (2005) Understanding the dynamics of thrombin in cardiovascular disease: Pathobiology and biochemistry for the clinician. Am Heart J 149(1 SUPPL.): S2-S8. Franchini M, Liumbruno GM, Bonfanti C & Lippi G (2016) The evolution of anticoagulant therapy. Blood Transfusion 14(2): Goodstadt L & Ponting CP (2004) Vitamin K epoxide reductase: Homology, active site and catalytic mechanism. Trends Biochem Sci 29(6): Granner D, Murray R & Rodwell V (2003) Harper s illustrated biochemistry 27 th edition USA Virginia VA Jin D-, Tie J- & Stafford DW (2007) The conversion of vitamin K epoxide to Vitamin K quinone and vitamin K quinone to vitamin K hydroquinone uses the same active site cysteines. Biochemistry 46(24): Koulu M & Tuomisto J (2000) Farmakologia ja toksikologia 6 th edition Suomi Kuopio Li W, Schulman S, Dutton RJ, Boyd D, Beckwith J & Rapoport TA (2010) Structure of a bacterial homologue of vitamin K epoxide reductase. Nature 463(7280): Mathews C, van Holde K & Ahern K (2008) Biochemistry 3 rd edition USA California CA Rishavy MA & Berkner KL (2008) Insight into the coupling mechanism of the vitamin K- dependent carboxylase: Mutation of histidine 160 disrupts glutamic acid carbanion 25
26 formation and efficient coupling of vitamin K epoxidation to glutamic acid carboxylation. Biochemistry 47(37): Rishavy MA, Hallgren KW, Wilson LA, Usubalieva A, Runge KW & Berkner KL (2013) The Vitamin K oxidoreductase is a multimer that efficiently reduces Vitamin K epoxide to hydroquinone to allow Vitamin K-dependent protein carboxylation. J Biol Chem 288(44): Rishavy MA, Usubalieva A, Hallgren KW & Berkner KL (2011) Novel insight into the mechanism of the vitamin K oxidoreductase (VKOR): Electron relay through Cys43 and Cys51 reduces VKOR to allow vitamin K reduction and facilitation of vitamin K- dependent protein carboxylation. J Biol Chem 286(9): Rost S, Fregin A, Ivaskevicius V, Conzelmann E, Hörtnagel K, Pelz H-, Lappegard K, Seifried E, Scharrer I, Tuddenham EGD, Müller CR, Strom TM & Oldenburg J (2004) Mutations in VKORC1 cause warfarin resistance and multiple coagulation factor deficiency type 2. Nature 427(6974): Schulman S, Wang B, Li W & Rapoport TA (2010) Vitamin K epoxide reductase prefers ER membrane-anchored thioredoxin-like redox partners. Proc Natl Acad Sci U S A 107(34): Shen G, Cui W, Zhang H, Zhou F, Huang W, Liu Q, Yang Y, Li S, Bowman GR, Sadler JE, Gross ML & Li W (2017) Warfarin traps human Vitamin K epoxide reductase in an intermediate state during electron transfer. Nat Struct Mol Biol 24(1): Tie J-, Jin D-, Straight DL & Stafford DW (2011) Functional study of the vitamin K cycle in mammalian cells. Blood 117(10): Tie J- & Stafford DW (2016) Structural and functional insights into enzymes of the vitamin K cycle. J Thromb Haemost 14(2): Van Horn WD (2013) Structural and functional insights into human vitamin K epoxide reductase and vitamin K epoxide reductase-like. Crit Rev Biochem Mol Biol 48(4): Voet D, Voet J & Pratt C (2008) Principles of biochemistry 3 rd edition USA New Jersey NJ 26
27 II LIITE 27
28 28
29 29
30 30
31 31
32 32
33 33
34 34
35 35
36 36
37 37
38 38
39 39
40 40
41 41
42 42
43 43
44 44
45 45
46 46
47 47
ENTSYYMIKATA- LYYSIN PERUSTEET (dos. Tuomas Haltia)
") ENTSYYMIKATA- LYYSIN PERUSTEET (dos. Tuomas Haltia) Elämän edellytykset: Solun täytyy pystyä (a) replikoitumaan (B) katalysoimaan tarvitsemiaan reaktioita tehokkaasti ja selektiivisesti eli sillä on oltava
ENTSYYMIKATA- LYYSIN PERUSTEET (dos. Tuomas Haltia) Elämän edellytykset: Solun täytyy pystyä (a) replikoitumaan (B) katalysoimaan tarvitsemiaan reaktioita tehokkaasti ja selektiivisesti eli sillä on oltava
Peptidi ---- F ----- K ----- V ----- R ----- H ----- A ---- A. Siirtäjä-RNA:n (trna:n) (3 ) AAG UUC CAC GCA GUG CGU (5 ) antikodonit
 (3 ) AAG UUC CAC GCA GUG CGU (5 ) antikodonit") Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 3 Pisteet / 20 Osa 1: Haluat selvittää -- F -- K -- V -- R -- H -- A peptidiä
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 3 Pisteet / 20 Osa 1: Haluat selvittää -- F -- K -- V -- R -- H -- A peptidiä
ELEC-C2210 Molekyyli- ja solubiologia
 ELEC-C2210 Molekyyli- ja solubiologia Entsyymikatalyysi Vuento & Heino ss. 66-75 ECB: Luku 3, s. 90-93 & luku 4, s. 144- Dos. Tuomas Haltia, Biotieteiden laitos, biokemia ja biotekniikka Miten entsyymit
ELEC-C2210 Molekyyli- ja solubiologia Entsyymikatalyysi Vuento & Heino ss. 66-75 ECB: Luku 3, s. 90-93 & luku 4, s. 144- Dos. Tuomas Haltia, Biotieteiden laitos, biokemia ja biotekniikka Miten entsyymit
Bioteknologian tutkinto-ohjelma Valintakoe Tehtävä 3 Pisteet / 30
 Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 3 Pisteet / 30 3. a) Alla on lyhyt jakso dsdna:ta, joka koodaa muutaman aminohappotähteen
Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 3 Pisteet / 30 3. a) Alla on lyhyt jakso dsdna:ta, joka koodaa muutaman aminohappotähteen
Peptidisynteesi. SPPS:n Periaate
 Tapio Nevalainen Lääkeainesynteesit II 2011 eptidisynteesi i eptidisynteesi Suoritetaan yleensä kiinteän faasin pinnalla; solid phase peptide synthesis (SS) Suuret peptidiainemäärät valmistetaan liuosfaasissa.
Tapio Nevalainen Lääkeainesynteesit II 2011 eptidisynteesi i eptidisynteesi Suoritetaan yleensä kiinteän faasin pinnalla; solid phase peptide synthesis (SS) Suuret peptidiainemäärät valmistetaan liuosfaasissa.
HYYTYMISVALMISTEIDEN HALLITTU KÄYTTÖ. Riitta Heino Erikoislääkäri TYKS, TOTEK
 HYYTYMISVALMISTEIDEN HALLITTU KÄYTTÖ Riitta Heino Erikoislääkäri TYKS, TOTEK ISCB 2009: KORVAUSPROTOKOLLAT 2010: FIBRINOGEENI 2011: HYYTYMISEN MONITOROINTI KORVAUKSEN HISTORIAA 70-luvulla: 2 litraa kirkkaita,
HYYTYMISVALMISTEIDEN HALLITTU KÄYTTÖ Riitta Heino Erikoislääkäri TYKS, TOTEK ISCB 2009: KORVAUSPROTOKOLLAT 2010: FIBRINOGEENI 2011: HYYTYMISEN MONITOROINTI KORVAUKSEN HISTORIAA 70-luvulla: 2 litraa kirkkaita,
Hyytymisjärjestelmä. hyytymistutkimukset
 VIIa Hyytymisjärjestelmä ja hyytymistutkimukset trombiini VIII VWF TF VIIa kudostekijää sisältävä solu TF VIIa Xa Va XIa Va trombosyytti VIIIa IXa VIIIa aktivoitunut trombosyytti Xa Va suuri määrä trombiinia
VIIa Hyytymisjärjestelmä ja hyytymistutkimukset trombiini VIII VWF TF VIIa kudostekijää sisältävä solu TF VIIa Xa Va XIa Va trombosyytti VIIIa IXa VIIIa aktivoitunut trombosyytti Xa Va suuri määrä trombiinia
Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe Tehtävä 1 Pisteet / 15
 Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
BI4 IHMISEN BIOLOGIA
 BI4 IHMISEN BIOLOGIA Verenkierto toimii elimistön kuljetusjärjestelmänä 6 Avainsanat fibriini fibrinogeeni hiussuoni hyytymistekijät imusuonisto iso verenkierto keuhkoverenkierto laskimo lepovaihe eli
BI4 IHMISEN BIOLOGIA Verenkierto toimii elimistön kuljetusjärjestelmänä 6 Avainsanat fibriini fibrinogeeni hiussuoni hyytymistekijät imusuonisto iso verenkierto keuhkoverenkierto laskimo lepovaihe eli
Biokemian perusteet 26.9.2012: Hemoglobiini, Entsyymikatalyysi
 Biokemian perusteet 26.9.2012: Hemoglobiini, Entsyymikatalyysi Dos. Tuomas Haltia Sirppisoluanemia, Hb-mutaatio Glu-6 Val Hemoglobiini allosteerinen hapen kuljettajaproteiini (ei ole entsyymi!) Allosteerinen
Biokemian perusteet 26.9.2012: Hemoglobiini, Entsyymikatalyysi Dos. Tuomas Haltia Sirppisoluanemia, Hb-mutaatio Glu-6 Val Hemoglobiini allosteerinen hapen kuljettajaproteiini (ei ole entsyymi!) Allosteerinen
Ma > GENERAL PRINCIPLES OF CELL SIGNALING
 Ma 5.12. -> GENERAL PRINCIPLES OF CELL SIGNALING Cell-Surface Receptors Relay Extracellular Signals via Intracellular Signaling Pathways Some Intracellular Signaling Proteins Act as Molecular Switches
Ma 5.12. -> GENERAL PRINCIPLES OF CELL SIGNALING Cell-Surface Receptors Relay Extracellular Signals via Intracellular Signaling Pathways Some Intracellular Signaling Proteins Act as Molecular Switches
PROTEIINIEN MUOKKAUS JA KULJETUS
 PROTEIINIEN MUOKKAUS JA KULJETUS 1.1 Endoplasmakalvosto Endoplasmakalvosto on organelli joka sijaitsee tumakalvossa kiinni. Se on topologisesti siis yhtä tumakotelon kanssa. Se koostuu kahdesta osasta:
PROTEIINIEN MUOKKAUS JA KULJETUS 1.1 Endoplasmakalvosto Endoplasmakalvosto on organelli joka sijaitsee tumakalvossa kiinni. Se on topologisesti siis yhtä tumakotelon kanssa. Se koostuu kahdesta osasta:
Vastaa lyhyesti selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1 1) Tunnista molekyylit (1 piste) ja täytä seuraava taulukko (2 pistettä) a) b) c) d) a) Syklinen AMP (camp) (0.25) b) Beta-karoteeni (0.25 p) c) Sakkaroosi (0.25 p) d) -D-Glukopyranoosi (0.25 p) 2 Taulukko.
1 1) Tunnista molekyylit (1 piste) ja täytä seuraava taulukko (2 pistettä) a) b) c) d) a) Syklinen AMP (camp) (0.25) b) Beta-karoteeni (0.25 p) c) Sakkaroosi (0.25 p) d) -D-Glukopyranoosi (0.25 p) 2 Taulukko.
Proteiinien muuntuminen energiaksi ihmiselimistössä
 Proteiinien muuntuminen energiaksi ihmiselimistössä Helsingin yliopisto Matemaattis-luonnontieteellinen tiedekunta Kemian laitos Kemian opettajankoulutusyksikkö Kandidaatintutkielma Tekijä: Klaus Sippel
Proteiinien muuntuminen energiaksi ihmiselimistössä Helsingin yliopisto Matemaattis-luonnontieteellinen tiedekunta Kemian laitos Kemian opettajankoulutusyksikkö Kandidaatintutkielma Tekijä: Klaus Sippel
Farmakogeneettiset testit apuna lääkehoidon arvioinnissa
 Farmakogeneettiset testit apuna lääkehoidon arvioinnissa Farmakogeneettiset testit Farmakogenetiikalla tarkoitetaan geneettisiä variaatioita, jotka vaikuttavat lääkeainevasteeseen. Geneettisen tiedon hyödyntäminen
Farmakogeneettiset testit apuna lääkehoidon arvioinnissa Farmakogeneettiset testit Farmakogenetiikalla tarkoitetaan geneettisiä variaatioita, jotka vaikuttavat lääkeainevasteeseen. Geneettisen tiedon hyödyntäminen
Henkilötunnus - Biokemian/bioteknologian valintakoe. Sukunimi Etunimet Tehtävä 1 Pisteet / 20
 elsingin yliopisto/tampereen yliopisto enkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24. 5. 2004 Etunimet Tehtävä 1 Pisteet / 20 Solujen kalvorakenteet rajaavat solut niiden ulkoisesta ympäristöstä
elsingin yliopisto/tampereen yliopisto enkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24. 5. 2004 Etunimet Tehtävä 1 Pisteet / 20 Solujen kalvorakenteet rajaavat solut niiden ulkoisesta ympäristöstä
OPPIMATERIAALI HYYTYMISJÄRJESTELMÄSTÄ JA HYYTYMISTUTKIMUKSISTA
 OPPIMATERIAALI HYYTYMISJÄRJESTELMÄSTÄ JA HYYTYMISTUTKIMUKSISTA Janita Hiltunen Karolina Laine Opinnäytetyö Lokakuu 2015 Bioanalyytikkokoulutus TIIVISTELMÄ Tampereen ammattikorkeakoulu Bioanalyytikkokoulutus
OPPIMATERIAALI HYYTYMISJÄRJESTELMÄSTÄ JA HYYTYMISTUTKIMUKSISTA Janita Hiltunen Karolina Laine Opinnäytetyö Lokakuu 2015 Bioanalyytikkokoulutus TIIVISTELMÄ Tampereen ammattikorkeakoulu Bioanalyytikkokoulutus
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
Mind Master. Matti Vire 11.5.2013
 Stressi = ympäristön yksilöön kohdistava uhka tai vahingollinen vaikutus sympaattinen hermojärjestelmä ja hypotalamus-aivolisäke-lisämunuainen aktivoituvat Akuutissa stressissä sydämen syke nousee, hengitys
Stressi = ympäristön yksilöön kohdistava uhka tai vahingollinen vaikutus sympaattinen hermojärjestelmä ja hypotalamus-aivolisäke-lisämunuainen aktivoituvat Akuutissa stressissä sydämen syke nousee, hengitys
Biomolekyylit 2. Nukleotidit, aminohapot ja proteiinit
 Biomolekyylit 2 Nukleotidit, aminohapot ja proteiinit Nukleotidit Ihmisen perimä, eli DNA (deoksiribonukleiinihappo) muodostuu pitkästä nukleotidiketjusta. Lisäksi nukleotidit toimivat mm. proteiinisynteesissä
Biomolekyylit 2 Nukleotidit, aminohapot ja proteiinit Nukleotidit Ihmisen perimä, eli DNA (deoksiribonukleiinihappo) muodostuu pitkästä nukleotidiketjusta. Lisäksi nukleotidit toimivat mm. proteiinisynteesissä
THROMBOTRACK SOLO HYYTYMISANALYSAATTO- RIN TYÖOHJE BIOANALY- TIIKAN OPISKELIJOILLE
 OPINNÄYTETYÖ - AMMATTIKORKEAKOULUTUTKINTO SOSIAALI-, TERVEYS- JA LIIKUNTA-ALA THROMBOTRACK SOLO HYYTYMISANALYSAATTO- RIN TYÖOHJE BIOANALY- TIIKAN OPISKELIJOILLE T E - K I J Ä / T : Leeni Tapiola SAVONIA-AMMATTIKORKEAKOULU
OPINNÄYTETYÖ - AMMATTIKORKEAKOULUTUTKINTO SOSIAALI-, TERVEYS- JA LIIKUNTA-ALA THROMBOTRACK SOLO HYYTYMISANALYSAATTO- RIN TYÖOHJE BIOANALY- TIIKAN OPISKELIJOILLE T E - K I J Ä / T : Leeni Tapiola SAVONIA-AMMATTIKORKEAKOULU
Leikkausverenvuodon portaittainen korvaus. Kati Järvelä TAYS Sydänkeskus Oy
 Leikkausverenvuodon portaittainen korvaus Kati Järvelä TAYS Sydänkeskus Oy Veritilavuus Hapenkuljetus kyky Hemostaasin ylläpito =verenvuodon tyrehdyttäminen / tyrehtyminen Veritilavuus Leikkausverenvuodon
Leikkausverenvuodon portaittainen korvaus Kati Järvelä TAYS Sydänkeskus Oy Veritilavuus Hapenkuljetus kyky Hemostaasin ylläpito =verenvuodon tyrehdyttäminen / tyrehtyminen Veritilavuus Leikkausverenvuodon
Broilerivehnän viljelypäivä 2.2.2012 Essi Tuomola
 Hankeaika 10.10.2007-31.12.2012 Yhteistyössä: Siipikarjan tuottajat Broilerivehnän viljelypäivä 2.2.2012 Essi Tuomola Broilerin rehustuksen koostumus Valkuaisaineet Aminohapot Vehnän rehuarvo broilerille
Hankeaika 10.10.2007-31.12.2012 Yhteistyössä: Siipikarjan tuottajat Broilerivehnän viljelypäivä 2.2.2012 Essi Tuomola Broilerin rehustuksen koostumus Valkuaisaineet Aminohapot Vehnän rehuarvo broilerille
Käsitteitä. Hormones and the Endocrine System Hormonit ja sisäeritejärjestelmä. Sisäeriterauhanen
 Käsitteitä Hormones and the Endocrine System Hormonit ja sisäeritejärjestelmä 1/2 Umpirauhanen vs. sisäeriterauhanen Endokrinologia Parakriininen Autokriininen Neurotransmitteri Reseptori Sisäeriterauhanen
Käsitteitä Hormones and the Endocrine System Hormonit ja sisäeritejärjestelmä 1/2 Umpirauhanen vs. sisäeriterauhanen Endokrinologia Parakriininen Autokriininen Neurotransmitteri Reseptori Sisäeriterauhanen
VASTAUS 1: Yhdistä oikein
 KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
7. MAKSA JA MUNUAISET
 7. MAKSA JA MUNUAISET 7.1. Maksa myrkkyjentuhoaja SIJAINTI: Vatsaontelon yläosassa, oikealla puolella, välittömästi pallean alla Painaa reilun kilon RAKENNE: KAKSI LOHKOA: VASEN JA OIKEA (suurempi), VÄLISSÄ
7. MAKSA JA MUNUAISET 7.1. Maksa myrkkyjentuhoaja SIJAINTI: Vatsaontelon yläosassa, oikealla puolella, välittömästi pallean alla Painaa reilun kilon RAKENNE: KAKSI LOHKOA: VASEN JA OIKEA (suurempi), VÄLISSÄ
Solun kemiallinen peruskoostumus eläinsolu. Solun kemia. Solun kemiallinen peruskoostumus bakteerisolu. Vesi 1
 Solun kemiallinen peruskoostumus eläinsolu Solun kemia paino-% Vesi 75-90 proteiinit 10-20 Lipidit 2 Hiilihydraatit 1 RNA/DNA 0,7/0,4 Epäorg. 1,5 Solun kemiallinen peruskoostumus bakteerisolu Vesi 1 paino-%
Solun kemiallinen peruskoostumus eläinsolu Solun kemia paino-% Vesi 75-90 proteiinit 10-20 Lipidit 2 Hiilihydraatit 1 RNA/DNA 0,7/0,4 Epäorg. 1,5 Solun kemiallinen peruskoostumus bakteerisolu Vesi 1 paino-%
8. Alkoholit, fenolit ja eetterit
 8. Alkoholit, fenolit ja eetterit SM -08 Alkoholit ovat orgaanisia yhdisteitä, joissa on yksi tai useampia -ryhmiä. Fenoleissa -ryhmä on kiinnittynyt aromaattiseen renkaaseen. Alkoholit voivat olla primäärisiä,
8. Alkoholit, fenolit ja eetterit SM -08 Alkoholit ovat orgaanisia yhdisteitä, joissa on yksi tai useampia -ryhmiä. Fenoleissa -ryhmä on kiinnittynyt aromaattiseen renkaaseen. Alkoholit voivat olla primäärisiä,
Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20
 Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20 3: Osa 1 Tumallisten solujen genomin toiminnassa sekä geenien
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20 3: Osa 1 Tumallisten solujen genomin toiminnassa sekä geenien
KEMIA HYVÄN VASTAUKSEN PIIRTEET
 BILÄÄKETIETEEN enkilötunnus: - KULUTUSJELMA Sukunimi: 20.5.2015 Etunimet: Nimikirjoitus: KEMIA Kuulustelu klo 9.00-13.00 YVÄN VASTAUKSEN PIIRTEET Tehtävämonisteen tehtäviin vastataan erilliselle vastausmonisteelle.
BILÄÄKETIETEEN enkilötunnus: - KULUTUSJELMA Sukunimi: 20.5.2015 Etunimet: Nimikirjoitus: KEMIA Kuulustelu klo 9.00-13.00 YVÄN VASTAUKSEN PIIRTEET Tehtävämonisteen tehtäviin vastataan erilliselle vastausmonisteelle.
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. Valitse listasta kunkin yhdisteen yleiskielessä käytettävä ei-systemaattinen nimi. (pisteet yht. 5p) a) C-vitamiini b) glukoosi c) etikkahappo d) salisyylihappo e) beta-karoteeni a. b. c. d. e. ksylitoli
1. Valitse listasta kunkin yhdisteen yleiskielessä käytettävä ei-systemaattinen nimi. (pisteet yht. 5p) a) C-vitamiini b) glukoosi c) etikkahappo d) salisyylihappo e) beta-karoteeni a. b. c. d. e. ksylitoli
NON-CODING RNA (ncrna)
") NON-CODING RNA (ncrna) 1. Yleistä NcRNA eli non-coding RNA tarkoittaa kaikkia proteiinia koodaamattomia rnamolekyylejä. Näistä yleisimmin tunnetut ovat ribosomaalinen RNA (rrna) sekä siirtäjä-rna (trna),
NON-CODING RNA (ncrna) 1. Yleistä NcRNA eli non-coding RNA tarkoittaa kaikkia proteiinia koodaamattomia rnamolekyylejä. Näistä yleisimmin tunnetut ovat ribosomaalinen RNA (rrna) sekä siirtäjä-rna (trna),
Vitamiinien puutostilat: Christian Ejkman (1858-1930) havaitsi ensimmäisenä vuonna 1888, että jonkin ravintotekijän puute aiheutti kanoilla
 havaitsi ensimmäisenä vuonna 1888, että jonkin ravintotekijän puute aiheutti kanoilla") Vitamiinit Vitamiinit ovat biologisesti aktiivisia orgaanisia yhdisteitä, joita elimistö tarvitsee, mutta ei itse pysty syntetisoimaan lainkaan tai riittävästi ja siksi niitä on saatava ravinnosta. Poikkeuksena
Vitamiinit Vitamiinit ovat biologisesti aktiivisia orgaanisia yhdisteitä, joita elimistö tarvitsee, mutta ei itse pysty syntetisoimaan lainkaan tai riittävästi ja siksi niitä on saatava ravinnosta. Poikkeuksena
Veren hyytymiseen vaikuttava lääkitys päivystyksessä
 Veren hyytymiseen vaikuttava lääkitys päivystyksessä Yleistä Veren hyytymiseen vaikuttavat lääkkeet ovat erittäin yleisiä Veren hyytymiseen vaikuttavat lääkkeet ovat yleensä potilaalle tarpeen Päivystysaikana
Veren hyytymiseen vaikuttava lääkitys päivystyksessä Yleistä Veren hyytymiseen vaikuttavat lääkkeet ovat erittäin yleisiä Veren hyytymiseen vaikuttavat lääkkeet ovat yleensä potilaalle tarpeen Päivystysaikana
Hemostaatit GKS koulutuspäivät 23-24.9.2010
 Päivi Pakarinen LKT Erikoislääkäri HYKS Naistenklinikka Hemostaatit GKS koulutuspäivät 23-24.9.2010 Sisältö Elimistön verenvuodonhallinta Mitä ovat Hemostaatit Missä tarvitaan Milloin harkitaan Miten valitaan
Päivi Pakarinen LKT Erikoislääkäri HYKS Naistenklinikka Hemostaatit GKS koulutuspäivät 23-24.9.2010 Sisältö Elimistön verenvuodonhallinta Mitä ovat Hemostaatit Missä tarvitaan Milloin harkitaan Miten valitaan
Clinical impact of serum proteins on drug delivery Felix Kratz, Bakheet Elsadek Journal of Controlled Release 161 (2012)
") Clinical impact of serum proteins on drug delivery Felix Kratz, Bakheet Elsadek Journal of Controlled Release 161 (2012) 429 445 Sampo Kurvonen 25.10.2017 Sisältö Plasmaproteiineista Albumiini Transferriini
Clinical impact of serum proteins on drug delivery Felix Kratz, Bakheet Elsadek Journal of Controlled Release 161 (2012) 429 445 Sampo Kurvonen 25.10.2017 Sisältö Plasmaproteiineista Albumiini Transferriini
Hyytymisjärjestelmän perustutkimukset. Tuukka Helin, LL Erikoistuva lääkäri, kl kemia Helsingin kaupunki, HUSLAB. Labquality-päivät 6.2.
 Hyytymisjärjestelmän perustutkimukset Tuukka Helin, LL Erikoistuva lääkäri, kl kemia Helsingin kaupunki, HUSLAB Labquality-päivät 6.2.2014 Sidonnaisuudet Työsuhteet Helsingin kaupunki, HUSLAB Matka-avustukset
Hyytymisjärjestelmän perustutkimukset Tuukka Helin, LL Erikoistuva lääkäri, kl kemia Helsingin kaupunki, HUSLAB Labquality-päivät 6.2.2014 Sidonnaisuudet Työsuhteet Helsingin kaupunki, HUSLAB Matka-avustukset
Hemostaasiongelmia päivystyspotilaalla. Sisätautilääkäripäivät 27.11.2009 LT Pirjo Mustonen
 Hemostaasiongelmia päivystyspotilaalla Sisätautilääkäripäivät 27.11.2009 LT Pirjo Mustonen 1 1 Esitelmän sisältö varfariinia käyttävä potilas, jolla on aivoverenvuoto maksan vajaatoimintapotilas, joka
Hemostaasiongelmia päivystyspotilaalla Sisätautilääkäripäivät 27.11.2009 LT Pirjo Mustonen 1 1 Esitelmän sisältö varfariinia käyttävä potilas, jolla on aivoverenvuoto maksan vajaatoimintapotilas, joka
Geriatripäivät 2013 Turku
 Eteisvärinäpotilaan antikoagulanttihoidon nykysuositukset Geriatripäivät 2013 Turku Matti Erkko OYL/Kardiologi TKS sydänpkl Normaali sinusrytmi ja eteisvärinä 2 2 Eteisvärinä on yleinen Eteisvärinä aiheuttaa
Eteisvärinäpotilaan antikoagulanttihoidon nykysuositukset Geriatripäivät 2013 Turku Matti Erkko OYL/Kardiologi TKS sydänpkl Normaali sinusrytmi ja eteisvärinä 2 2 Eteisvärinä on yleinen Eteisvärinä aiheuttaa
NITRIFIKAATIOBAKTEERIEN TOIMINTA
 NITRIFIKAATIOBAKTEERIEN TOIMINTA 1(6) Ville Kivisalmi Typen kiertoon maa- ja vesiekosysteemeissä osallistuvat bakteerit ovat pääasiassa autotrofeja kemolitotrofeja, jotka saavat energiansa epäorgaanisten
NITRIFIKAATIOBAKTEERIEN TOIMINTA 1(6) Ville Kivisalmi Typen kiertoon maa- ja vesiekosysteemeissä osallistuvat bakteerit ovat pääasiassa autotrofeja kemolitotrofeja, jotka saavat energiansa epäorgaanisten
High Definition Body Lift selluliittigeeli
 High Definition Body Lift selluliittigeeli Lehdistötiedote helmikuu 2009 Paras tapa huolehtia vartalon virtaviivaisesta ulkonäöstä on syödä terveellisesti ja liikkua säännöllisesti. Liikunta ja runsaasti
High Definition Body Lift selluliittigeeli Lehdistötiedote helmikuu 2009 Paras tapa huolehtia vartalon virtaviivaisesta ulkonäöstä on syödä terveellisesti ja liikkua säännöllisesti. Liikunta ja runsaasti
Syöpä. Ihmisen keho muodostuu miljardeista soluista. Vaikka. EGF-kasvutekijä. reseptori. tuma. dna
 Ihmisen keho muodostuu miljardeista soluista. Vaikka nämä solut ovat tietyssä mielessä meidän omiamme, ne polveutuvat itsenäisistä yksisoluisista elämänmuodoista, jotka ovat säilyttäneet monia itsenäisen
Ihmisen keho muodostuu miljardeista soluista. Vaikka nämä solut ovat tietyssä mielessä meidän omiamme, ne polveutuvat itsenäisistä yksisoluisista elämänmuodoista, jotka ovat säilyttäneet monia itsenäisen
KandiakatemiA Kandiklinikka
 Kandiklinikka Kandit vastaavat Immunologia Luonnollinen ja hankittu immuniteetti IMMUNOLOGIA Ihmisen immuniteetti pohjautuu luonnolliseen ja hankittuun immuniteettiin. Immunologiasta vastaa lymfaattiset
Kandiklinikka Kandit vastaavat Immunologia Luonnollinen ja hankittu immuniteetti IMMUNOLOGIA Ihmisen immuniteetti pohjautuu luonnolliseen ja hankittuun immuniteettiin. Immunologiasta vastaa lymfaattiset
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi
 6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
INR MITTAUSTAVAT: PERINTEINEN LABORATORIOMITTAUS JA PIKAMITTAUS (VIERITESTAUS) 30.3.2016
 30.3.2016") INR MITTAUSTAVAT: PERINTEINEN LABORATORIOMITTAUS JA PIKAMITTAUS (VIERITESTAUS) 30.3.2016 Elina Porkkala-Sarataho Tuotantopäällikkö Fimlab Laboratoriot Oy Keski-Suomen alue SISÄLTÖ Tromboplastiiniaika,
INR MITTAUSTAVAT: PERINTEINEN LABORATORIOMITTAUS JA PIKAMITTAUS (VIERITESTAUS) 30.3.2016 Elina Porkkala-Sarataho Tuotantopäällikkö Fimlab Laboratoriot Oy Keski-Suomen alue SISÄLTÖ Tromboplastiiniaika,
DNA RNA proteiinit transkriptio prosessointi translaatio regulaatio
 CELL 411-- replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi
CELL 411-- replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi
Vitamiinit. Tärkeimpiä lähteitä: maksa, maitotuotteet, porkkana, parsakaali ja pinaatti
 Vitamiinit A-vitamiini Tärkeimpiä lähteitä: maksa, maitotuotteet, porkkana, parsakaali ja pinaatti Välttämätön näköaistimuksen syntyyn hämärässä, solujen kasvuun ja erilaistumiseen sekä ihmisen lisääntymiseen.
Vitamiinit A-vitamiini Tärkeimpiä lähteitä: maksa, maitotuotteet, porkkana, parsakaali ja pinaatti Välttämätön näköaistimuksen syntyyn hämärässä, solujen kasvuun ja erilaistumiseen sekä ihmisen lisääntymiseen.
Fimea kehittää, arvioi ja informoi
 Fimea kehittää, arvioi ja informoi SELKOTIIVISTELMÄ JULKAISUSARJA 4/2012 Eteisvärinän hoito Verenohennuslääke dabigatraanin ja varfariinin vertailu Eteisvärinä on sydämen rytmihäiriö, joka voi aiheuttaa
Fimea kehittää, arvioi ja informoi SELKOTIIVISTELMÄ JULKAISUSARJA 4/2012 Eteisvärinän hoito Verenohennuslääke dabigatraanin ja varfariinin vertailu Eteisvärinä on sydämen rytmihäiriö, joka voi aiheuttaa
PAKASTETUN P-APTT-NÄYTTEEN SÄILYVYYS AJAN FUNKTIONA
 PAKASTETUN P-APTT-NÄYTTEEN SÄILYVYYS AJAN FUNKTIONA Terhi Anttila Outi Halttu Opinnäytetyö Lokakuu 2015 Bioanalyytikko-koulutus TIIVISTELMÄ Tampereen ammattikorkeakoulu Bioanalyytikko-koulutus 12BIO ANTTILA,
PAKASTETUN P-APTT-NÄYTTEEN SÄILYVYYS AJAN FUNKTIONA Terhi Anttila Outi Halttu Opinnäytetyö Lokakuu 2015 Bioanalyytikko-koulutus TIIVISTELMÄ Tampereen ammattikorkeakoulu Bioanalyytikko-koulutus 12BIO ANTTILA,
KEMIA. Kemia on tiede joka tutkii aineen koostumuksia, ominaisuuksia ja muuttumista.
 KEMIA Kemia on tiede joka tutkii aineen koostumuksia, ominaisuuksia ja muuttumista. Kemian työturvallisuudesta -Kemian tunneilla tutustutaan aineiden ominaisuuksiin Jotkin aineet syttyvät palamaan reagoidessaan
KEMIA Kemia on tiede joka tutkii aineen koostumuksia, ominaisuuksia ja muuttumista. Kemian työturvallisuudesta -Kemian tunneilla tutustutaan aineiden ominaisuuksiin Jotkin aineet syttyvät palamaan reagoidessaan
Toimenpiteeseen tulevan potilaan antikoagulaatiohoito ja veren hyytymishäiriöt. Jarkko Karihuhta 9.10.2014
 Toimenpiteeseen tulevan potilaan antikoagulaatiohoito ja veren hyytymishäiriöt Jarkko Karihuhta 9.10.2014 Hemostaasi Verenhyytymisjärjestelmässä Verihiutaleet Reagoivat verisuonen sisäpinnan endoteelivaurioon
Toimenpiteeseen tulevan potilaan antikoagulaatiohoito ja veren hyytymishäiriöt Jarkko Karihuhta 9.10.2014 Hemostaasi Verenhyytymisjärjestelmässä Verihiutaleet Reagoivat verisuonen sisäpinnan endoteelivaurioon
ARGATROBAANI (NOVASTAN ) INFUUSIO - OHJE
 INFUUSIO - OHJE") ARGATROBAANI (NOVASTAN ) INFUUSIO - OHJE Argatrobaani (Novastan ) on suora trombiinin estäjä, joka sitoutuu palautuvasti trombiiniin. Argatrobaani pystyy estämään sekä vapaan että hyytymiin kiinnittyneen
ARGATROBAANI (NOVASTAN ) INFUUSIO - OHJE Argatrobaani (Novastan ) on suora trombiinin estäjä, joka sitoutuu palautuvasti trombiiniin. Argatrobaani pystyy estämään sekä vapaan että hyytymiin kiinnittyneen
Proteiinin rakenteen selvittämisestä ja visualisoinnista
 TKK Solubiosysteemien perusteet syksy 2002 Harkkatyö M.Tarvainen Proteiinin rakenteen selvittämisestä ja visualisoinnista 1. Yleistä proteiineista 2. Röntgensädekristallografia 3. Ydinmagneettinen resonanssimenetelmä
TKK Solubiosysteemien perusteet syksy 2002 Harkkatyö M.Tarvainen Proteiinin rakenteen selvittämisestä ja visualisoinnista 1. Yleistä proteiineista 2. Röntgensädekristallografia 3. Ydinmagneettinen resonanssimenetelmä
Uutta antikoagulaatiosta: onko marevan mennyttä? Anne Pinomäki, LL Osastonlääkäri Hyytymishäiriöyksikkö BioChem 24.3.2011
 Uutta antikoagulaatiosta: onko marevan mennyttä? Anne Pinomäki, LL Osastonlääkäri Hyytymishäiriöyksikkö BioChem 24.3.2011 Johdanto Yli 2 % väestöstä antikoagulaatiohoidon piirissä Useamman viime vuoden
Uutta antikoagulaatiosta: onko marevan mennyttä? Anne Pinomäki, LL Osastonlääkäri Hyytymishäiriöyksikkö BioChem 24.3.2011 Johdanto Yli 2 % väestöstä antikoagulaatiohoidon piirissä Useamman viime vuoden
Liite III. Muutokset valmisteyhteenvedon ja pakkausselosteiden asianmukaisiin kohtiin
 Liite III Muutokset valmisteyhteenvedon ja pakkausselosteiden asianmukaisiin kohtiin Huomautus: Seuraavat muutokset valmisteyhteenvedon ja pakkausselosteen tiettyihin kohtiin tehdään sovittelumenettelyn
Liite III Muutokset valmisteyhteenvedon ja pakkausselosteiden asianmukaisiin kohtiin Huomautus: Seuraavat muutokset valmisteyhteenvedon ja pakkausselosteen tiettyihin kohtiin tehdään sovittelumenettelyn
P-TT-INR-NÄYTTEEN SÄILYVYYS KOKOVERENÄ NÄYTTEENOTTO- PUTKESSA HUONEENLÄMMÖSSÄ
 P-TT-INR-NÄYTTEEN SÄILYVYYS KOKOVERENÄ NÄYTTEENOTTO- PUTKESSA HUONEENLÄMMÖSSÄ Maija Heiska Erika Liimatainen Opinnäytetyö Syyskuu 2015 Bioanalyytikkokoulutus TIIVISTELMÄ Tampereen ammattikorkeakoulu Bioanalyytikkokoulutus
P-TT-INR-NÄYTTEEN SÄILYVYYS KOKOVERENÄ NÄYTTEENOTTO- PUTKESSA HUONEENLÄMMÖSSÄ Maija Heiska Erika Liimatainen Opinnäytetyö Syyskuu 2015 Bioanalyytikkokoulutus TIIVISTELMÄ Tampereen ammattikorkeakoulu Bioanalyytikkokoulutus
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita
 Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 10. Valkuaisaineiden valmistaminen solussa 1. Avainsanat 2. Perinnöllinen tieto on dna:n emäsjärjestyksessä 3. Proteiinit koostuvat
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 10. Valkuaisaineiden valmistaminen solussa 1. Avainsanat 2. Perinnöllinen tieto on dna:n emäsjärjestyksessä 3. Proteiinit koostuvat
Entsyymit ja niiden tuotanto. Niklas von Weymarn, VTT Erikoistutkija ja tiiminvetäjä
 Entsyymit ja niiden tuotanto Niklas von Weymarn, VTT Erikoistutkija ja tiiminvetäjä Mitä ovat entsyymit? Entsyymit ovat proteiineja (eli valkuaisaineita), jotka vauhdittavat (katalysoivat) kemiallisia
Entsyymit ja niiden tuotanto Niklas von Weymarn, VTT Erikoistutkija ja tiiminvetäjä Mitä ovat entsyymit? Entsyymit ovat proteiineja (eli valkuaisaineita), jotka vauhdittavat (katalysoivat) kemiallisia
Veren hyytymistapahtuma ja rekombinantti FVIIa
 Veren hyytymistapahtuma ja rekombinantti FVIIa Veli-Matti Karjalainen Jopa neljännes A-hemofi ilikoista kehittää vasta-aineita tekijälle VIII. Hoitona on käytetty joko valtavan isoja FVIII annoksia tai
Veren hyytymistapahtuma ja rekombinantti FVIIa Veli-Matti Karjalainen Jopa neljännes A-hemofi ilikoista kehittää vasta-aineita tekijälle VIII. Hoitona on käytetty joko valtavan isoja FVIII annoksia tai
Kertausluennot: Mahdollisuus pisteiden korotukseen ja rästisuorituksiin Keskiviikko klo 8-10
 Kertausluennot: Mahdollisuus pisteiden korotukseen ja rästisuorituksiin Keskiviikko 25.10 klo 8-10 Jokaisesta oikein ratkaistusta tehtävästä voi saada yhden lisäpisteen. Tehtävä, joilla voi korottaa kotitehtävän
Kertausluennot: Mahdollisuus pisteiden korotukseen ja rästisuorituksiin Keskiviikko 25.10 klo 8-10 Jokaisesta oikein ratkaistusta tehtävästä voi saada yhden lisäpisteen. Tehtävä, joilla voi korottaa kotitehtävän
Solun perusrakenne I Solun perusrakenne. BI2 I Solun perusrakenne 3. Solujen kemiallinen rakenne
 Solun perusrakenne I Solun perusrakenne 3. Solujen kemiallinen rakenne 1. Avainsanat 2. Solut koostuvat molekyyleistä 3. Hiilihydraatit 4. Lipidit eli rasva-aineet 5. Valkuaisaineet eli proteiinit rakentuvat
Solun perusrakenne I Solun perusrakenne 3. Solujen kemiallinen rakenne 1. Avainsanat 2. Solut koostuvat molekyyleistä 3. Hiilihydraatit 4. Lipidit eli rasva-aineet 5. Valkuaisaineet eli proteiinit rakentuvat
Laskuharjoitus 3 palautus 11. 11. 2003 mennessä. Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa):
:") Laskuharjoitus 3 palautus 11. 11. 2003 mennessä Tehtävä 1: Entsyymikinetiikkaa Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa): 1. A:n sitoutuminen saa konformaatiossa aikaan muutoksen,
Laskuharjoitus 3 palautus 11. 11. 2003 mennessä Tehtävä 1: Entsyymikinetiikkaa Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa): 1. A:n sitoutuminen saa konformaatiossa aikaan muutoksen,
CHEM-A1200 Kemiallinen rakenne ja sitoutuminen
 CHEM-A1200 Kemiallinen rakenne ja sitoutuminen Hapot, Emäkset ja pk a Opettava tutkija Pekka M Joensuu Jokaisella hapolla on: Arvo, joka kertoo meille kuinka hapan kyseinen protoni on. Helpottaa valitsemaan
CHEM-A1200 Kemiallinen rakenne ja sitoutuminen Hapot, Emäkset ja pk a Opettava tutkija Pekka M Joensuu Jokaisella hapolla on: Arvo, joka kertoo meille kuinka hapan kyseinen protoni on. Helpottaa valitsemaan
Itseopiskelumateriaali hyytymistekijöistä ja niiden laboratoriotutkimuksista
 Henna Kreander Itseopiskelumateriaali hyytymistekijöistä ja niiden laboratoriotutkimuksista Kliininen hematologia Metropolia Ammattikorkeakoulu Bioanalyytikko (AMK) Bioanalyytikon koulutusohjelma Opinnäytetyö
Henna Kreander Itseopiskelumateriaali hyytymistekijöistä ja niiden laboratoriotutkimuksista Kliininen hematologia Metropolia Ammattikorkeakoulu Bioanalyytikko (AMK) Bioanalyytikon koulutusohjelma Opinnäytetyö
Biomolekyylit ja biomeerit
 Biomolekyylit ja biomeerit Polymeerit ovat hyvin suurikokoisia, pitkäketjuisia molekyylejä, jotka muodostuvat monomeereista joko polyadditio- tai polykondensaatioreaktiolla. Polymeerit Synteettiset polymeerit
Biomolekyylit ja biomeerit Polymeerit ovat hyvin suurikokoisia, pitkäketjuisia molekyylejä, jotka muodostuvat monomeereista joko polyadditio- tai polykondensaatioreaktiolla. Polymeerit Synteettiset polymeerit
Opas laboratoriohenkilökunnalle
 Opas laboratoriohenkilökunnalle HEMLIBRA (emisitsumabi) Tähän lääkkeeseen kohdistuu lisäseuranta. Tällä tavalla voidaan havaita nopeasti uutta turvallisuutta koskevaa tietoa. Voit auttaa ilmoittamalla
Opas laboratoriohenkilökunnalle HEMLIBRA (emisitsumabi) Tähän lääkkeeseen kohdistuu lisäseuranta. Tällä tavalla voidaan havaita nopeasti uutta turvallisuutta koskevaa tietoa. Voit auttaa ilmoittamalla
Peptidisynteesi. Kiinteän faasin peptidisynteesi. SPPS:n Periaate Aminohappo kytketää. synthesis (SPPS) Suuret peptidiainemää
 Suuret peptidiainemää") eptidisynteesi uoritetaan yleensä kiinteän n faasin pinnalla; solid phase peptide synthesis () uuret peptidiainemää äärät t valmistetaan liuosfaasissa.. B. Merrifield: ribonukleaasientsyymin synteesi vuonna
eptidisynteesi uoritetaan yleensä kiinteän n faasin pinnalla; solid phase peptide synthesis () uuret peptidiainemää äärät t valmistetaan liuosfaasissa.. B. Merrifield: ribonukleaasientsyymin synteesi vuonna
Neuropeptidit, opiaatit ja niihin liittyvät mekanismit. Pertti Panula Biolääketieteen laitos 2013
 Neuropeptidit, opiaatit ja niihin liittyvät mekanismit Pertti Panula Biolääketieteen laitos 2013 Neuroendokriinisen järjestelmän säätely elimistössä Neuropeptidit Peptidirakenteisia hermovälittäjäaineita
Neuropeptidit, opiaatit ja niihin liittyvät mekanismit Pertti Panula Biolääketieteen laitos 2013 Neuroendokriinisen järjestelmän säätely elimistössä Neuropeptidit Peptidirakenteisia hermovälittäjäaineita
The Plant Cell / Fotosynteesi
 The Plant Cell / Fotosynteesi Solubiologian luennot 2003, kasvitiede Valoreaktiot Klorofyllimolekyylit ja muut pigmenttiaineet etenkin karotenoidit muodostavat valohaavin (200-300 pigmenttimolekyyliä),
The Plant Cell / Fotosynteesi Solubiologian luennot 2003, kasvitiede Valoreaktiot Klorofyllimolekyylit ja muut pigmenttiaineet etenkin karotenoidit muodostavat valohaavin (200-300 pigmenttimolekyyliä),
VALMISTEYHTEENVETO 1. LÄÄKEVALMISTEEN NIMI. Octaplex, 500 IU, infuusiokuiva-aine ja liuotin, liuosta varten 2. VAIKUTTAVAT AINEET JA NIIDEN MÄÄRÄT
 VALMISTEYHTEENVETO 1. LÄÄKEVALMISTEEN NIMI Octaplex, 500 IU, infuusiokuiva-aine ja liuotin, liuosta varten 2. VAIKUTTAVAT AINEET JA NIIDEN MÄÄRÄT Octaplex-valmiste koostuu ihmisen protrombiinikompleksia
VALMISTEYHTEENVETO 1. LÄÄKEVALMISTEEN NIMI Octaplex, 500 IU, infuusiokuiva-aine ja liuotin, liuosta varten 2. VAIKUTTAVAT AINEET JA NIIDEN MÄÄRÄT Octaplex-valmiste koostuu ihmisen protrombiinikompleksia
Anatomia ja fysiologia 1
 Anatomia ja fysiologia 1 Tehtävät Laura Partanen 2 Sisällysluettelo Solu... 3 Aktiopotentiaali... 4 Synapsi... 5 Iho... 6 Elimistön kemiallinen koostumus... 7 Kudokset... 8 Veri... 9 Sydän... 10 EKG...
Anatomia ja fysiologia 1 Tehtävät Laura Partanen 2 Sisällysluettelo Solu... 3 Aktiopotentiaali... 4 Synapsi... 5 Iho... 6 Elimistön kemiallinen koostumus... 7 Kudokset... 8 Veri... 9 Sydän... 10 EKG...
Kananmunatutkimusta suomalaisessa väestötutkimuksessa
 Kananmunatutkimusta suomalaisessa väestötutkimuksessa Helsinki, 27.11.2018 Jyrki Virtanen, FT, ravitsemusepidemiologian dosentti Laillistettu ravitsemusterapeutti Esityksen sisältö Kananmunan ravitsemuksellinen
Kananmunatutkimusta suomalaisessa väestötutkimuksessa Helsinki, 27.11.2018 Jyrki Virtanen, FT, ravitsemusepidemiologian dosentti Laillistettu ravitsemusterapeutti Esityksen sisältö Kananmunan ravitsemuksellinen
Laskuharjoitus 2 vastauksia ja selityksiä
 Laskuharjoitus vastauksia ja selityksiä Selitykset ovat usein jonkin verran laajempia kuin vastaukseen edellytetyt, jotta asia selviäisi niillekin, jotka eivät sitä aluksi olleet keksineet. Tehtävä : Proteiinien
Laskuharjoitus vastauksia ja selityksiä Selitykset ovat usein jonkin verran laajempia kuin vastaukseen edellytetyt, jotta asia selviäisi niillekin, jotka eivät sitä aluksi olleet keksineet. Tehtävä : Proteiinien
BI4 IHMISEN BIOLOGIA
 BI4 IHMISEN BIOLOGIA IHMINEN ON TOIMIVA KOKONAISUUS Ihmisessä on noin 60 000 miljardia solua Solujen perusrakenne on samanlainen, mutta ne ovat erilaistuneet hoitamaan omia tehtäviään Solujen on oltava
BI4 IHMISEN BIOLOGIA IHMINEN ON TOIMIVA KOKONAISUUS Ihmisessä on noin 60 000 miljardia solua Solujen perusrakenne on samanlainen, mutta ne ovat erilaistuneet hoitamaan omia tehtäviään Solujen on oltava
Kaikenlaisia sidoksia yhdisteissä: ioni-, kovalenttiset ja metallisidokset Fysiikan ja kemian perusteet ja pedagogiikka
 Kaikenlaisia sidoksia yhdisteissä: ioni-, kovalenttiset ja metallisidokset Fysiikan ja kemian perusteet ja pedagogiikka Kari Sormunen Kevät 2012 Kertausta IONIEN MUODOSTUMISESTA Jos atomi luovuttaa tai
Kaikenlaisia sidoksia yhdisteissä: ioni-, kovalenttiset ja metallisidokset Fysiikan ja kemian perusteet ja pedagogiikka Kari Sormunen Kevät 2012 Kertausta IONIEN MUODOSTUMISESTA Jos atomi luovuttaa tai
Sukunimi 26. 05. 2005 Etunimet Tehtävä 1 Pisteet / 20
 elsingin yliopisto/tampereen yliopisto enkilötunnus - Biokemian/bioteknologian valintakoe ukunimi 26. 05. 2005 Etunimet Tehtävä 1 Pisteet / 20 olujen kalvorakenteiden perusrakenteen muodostavat amfipaattiset
elsingin yliopisto/tampereen yliopisto enkilötunnus - Biokemian/bioteknologian valintakoe ukunimi 26. 05. 2005 Etunimet Tehtävä 1 Pisteet / 20 olujen kalvorakenteiden perusrakenteen muodostavat amfipaattiset
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Etunimet Tehtävä 5 Pisteet / 20
 Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 5 Pisteet / 20 Glukoosidehydrogenaasientsyymi katalysoi glukoosin oksidaatiota
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 5 Pisteet / 20 Glukoosidehydrogenaasientsyymi katalysoi glukoosin oksidaatiota
Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Tuomas Haltia Elämälle (solulle) välttämättömiä asioita ovat:
 välttämättömiä asioita ovat:") Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Tuomas Haltia 3.12.2012 Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien
Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Tuomas Haltia 3.12.2012 Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien
Laskuharjoitus 3 palautus 11. 11. 2003 mennessä
 Laskuharjoitus 3 palautus 11. 11. 23 mennessä Tehtävä 1: Entsyymikinetiikkaa Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa): 1. A:n sitoutuminen saa konformaatiossa aikaan muutoksen,
Laskuharjoitus 3 palautus 11. 11. 23 mennessä Tehtävä 1: Entsyymikinetiikkaa Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa): 1. A:n sitoutuminen saa konformaatiossa aikaan muutoksen,
Solun Kalvot. Kalvot muodostuvat spontaanisti. Biologiset kalvot koostuvat tuhansista erilaisista molekyyleistä
 Solun Kalvot (ja Mallikalvot) Biologiset kalvot koostuvat tuhansista erilaisista molekyyleistä Biokemian ja Farmakologian erusteet 2012 Kalvot muodostuvat spontaanisti Veden rakenne => ydrofobinen vuorovaikutus
Solun Kalvot (ja Mallikalvot) Biologiset kalvot koostuvat tuhansista erilaisista molekyyleistä Biokemian ja Farmakologian erusteet 2012 Kalvot muodostuvat spontaanisti Veden rakenne => ydrofobinen vuorovaikutus
Hyötyosuus. ANNOS ja sen merkitys lääkehoidossa? Farmakokinetiikan perusteita. Solukalvon läpäisy. Alkureitin metabolia
 Neurofarmakologia Farmakologian perusteiden kertausta Pekka Rauhala Syksy 2012 Koulu, Mervaala & Tuomisto, 8. painos, 2012 Kappaleet 11-30 (pois kappale 18) tai vastaavat asiat muista oppikirjoista n.
Neurofarmakologia Farmakologian perusteiden kertausta Pekka Rauhala Syksy 2012 Koulu, Mervaala & Tuomisto, 8. painos, 2012 Kappaleet 11-30 (pois kappale 18) tai vastaavat asiat muista oppikirjoista n.
OHJEET SYDÄMEN JA VERISUONIEN TERVEYTEEN. Kuopiolaistutkimus: Sydänperäinen äkkikuolema kaataa myös nuoria ja kovakuntoisia
 OHJEET SYDÄMEN JA VERISUONIEN TERVEYTEEN Kuopiolaistutkimus: Sydänperäinen äkkikuolema kaataa myös nuoria ja kovakuntoisia Kymmenvaiheinen ohjelma sepelvaltimoiden terveyden ylläpitoon luontaishoidoilla
OHJEET SYDÄMEN JA VERISUONIEN TERVEYTEEN Kuopiolaistutkimus: Sydänperäinen äkkikuolema kaataa myös nuoria ja kovakuntoisia Kymmenvaiheinen ohjelma sepelvaltimoiden terveyden ylläpitoon luontaishoidoilla
Kissa: Leikkauksen jälkeisen kivun lievitys kohdun ja munasarjojen poistoleikkauksen sekä pienten pehmytkudoskirurgisten toimenpiteiden jälkeen.
 1. ELÄINLÄÄKKEEN NIMI Acticam 5 mg/ml injektioneste, liuos koirille ja kissoille 2. LAADULLINEN JA MÄÄRÄLLINEN KOOSTUMUS Yksi ml Acticam 5 mg/ml injektionesteliuosta sisältää: Vaikuttavat aineet Meloksikaami
1. ELÄINLÄÄKKEEN NIMI Acticam 5 mg/ml injektioneste, liuos koirille ja kissoille 2. LAADULLINEN JA MÄÄRÄLLINEN KOOSTUMUS Yksi ml Acticam 5 mg/ml injektionesteliuosta sisältää: Vaikuttavat aineet Meloksikaami
ULKOELEKTRONIRAKENNE JA METALLILUONNE
 ULKOELEKTRONIRAKENNE JA METALLILUONNE Palautetaan mieleen jaksollinen järjestelmä ja mitä siitä saa- Kertausta daan irti. H RYHMÄT OVAT SARAKKEITA Mitä sarakkeen numero kertoo? JAKSOT OVAT RIVEJÄ Mitä
ULKOELEKTRONIRAKENNE JA METALLILUONNE Palautetaan mieleen jaksollinen järjestelmä ja mitä siitä saa- Kertausta daan irti. H RYHMÄT OVAT SARAKKEITA Mitä sarakkeen numero kertoo? JAKSOT OVAT RIVEJÄ Mitä
Aminohapot ja proteiinit
 Aminohapot ja proteiinit Proteiinit ovat aminohappoketjusta muodostuvia ihmiselle välttämättömiä yhdisteitä tai useammasta aminohappoketjusta muodostuvia komplekseja. Lähes kaikilla tunnetuilla eliöillä
Aminohapot ja proteiinit Proteiinit ovat aminohappoketjusta muodostuvia ihmiselle välttämättömiä yhdisteitä tai useammasta aminohappoketjusta muodostuvia komplekseja. Lähes kaikilla tunnetuilla eliöillä
Massiivinen verenvuoto. Jyväskylä 4.10.2012 Markku Leh9mäki K- SKS
 Massiivinen verenvuoto Jyväskylä 4.10.2012 Markku Leh9mäki K- SKS Sisältö Määritelmä Veren fysiologiaa Vuodoilla on eroja Trauma Obstetrinen Massiivivuodon hoidosta Osatekijät Laboratoriomonitoroin9 K-
Massiivinen verenvuoto Jyväskylä 4.10.2012 Markku Leh9mäki K- SKS Sisältö Määritelmä Veren fysiologiaa Vuodoilla on eroja Trauma Obstetrinen Massiivivuodon hoidosta Osatekijät Laboratoriomonitoroin9 K-
Valtuuskunnille toimitetaan oheisena asiakirja C(2017) 3664 final LIITTEET 1 2.
 3664 final LIITTEET 1 2.") Euroopan unionin neuvosto Bryssel, 6. kesäkuuta 2017 (OR. en) 10021/17 ADD 1 SAATE Lähettäjä: Saapunut: 2. kesäkuuta 2017 Vastaanottaja: DENLEG 47 AGRI 309 SAN 242 DELACT 91 Euroopan komission pääsihteerin
Euroopan unionin neuvosto Bryssel, 6. kesäkuuta 2017 (OR. en) 10021/17 ADD 1 SAATE Lähettäjä: Saapunut: 2. kesäkuuta 2017 Vastaanottaja: DENLEG 47 AGRI 309 SAN 242 DELACT 91 Euroopan komission pääsihteerin
Oksidatiivinen fosforylaatio = ATP:n tuotto NADH:lta ja FADH2:lta hapelle tapahtuvan elektroninsiirron ja ATP-syntaasin avulla
 Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien elektronien vastaanottajana
Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien elektronien vastaanottajana
K1- ja K2-vitamiinien terveysvaikutukset
 K1- ja K2-vitamiinien terveysvaikutukset K1- ja K2-vitamiinien terveysvaikutukset ovat kasvavan mielenkiinnon kohteena. K- vitamiineja pidetään elimistön hyvinvoinnille erittäin tärkeinä, mutta yleisesti
K1- ja K2-vitamiinien terveysvaikutukset K1- ja K2-vitamiinien terveysvaikutukset ovat kasvavan mielenkiinnon kohteena. K- vitamiineja pidetään elimistön hyvinvoinnille erittäin tärkeinä, mutta yleisesti
Hyytymishäiriöt - laboratoriotutkimukset
 Hyytymishäiriöt - laboratoriotutkimukset Lotta Joutsi-Korhonen LT, dos, kliinisen kemian erikoislääkäri 4.12.2012 Kliininen kemia Helsingin yliopisto ja HUSLAB Hyytymishäiriöpotilas klinikassa Kliiniset
Hyytymishäiriöt - laboratoriotutkimukset Lotta Joutsi-Korhonen LT, dos, kliinisen kemian erikoislääkäri 4.12.2012 Kliininen kemia Helsingin yliopisto ja HUSLAB Hyytymishäiriöpotilas klinikassa Kliiniset
Kysymyksiä ja vastauksia
 PIKAOPAS Kysymyksiä ja vastauksia K: Mikä ja mihin käyttötarkoitukseen Acetium on? V: Acetium on tarkoitettu karsinogeenisen asetaldehydin sitomiseen paikallisesti mahasta ihmisillä, jotka kuuluvat asetaldehydialtistuksen
PIKAOPAS Kysymyksiä ja vastauksia K: Mikä ja mihin käyttötarkoitukseen Acetium on? V: Acetium on tarkoitettu karsinogeenisen asetaldehydin sitomiseen paikallisesti mahasta ihmisillä, jotka kuuluvat asetaldehydialtistuksen
sosiaaliturvatunnus Tehtävissä tarvittavia atomipainoja: hiili 12,01; vety 1,008; happi 16,00. Toisen asteen yhtälön ratkaisukaava: ax 2 + bx + c = 0;
 Valintakoe 2012 / Biokemia Nimi sosiaaliturvatunnus Tehtävissä tarvittavia atomipainoja: hiili 12,01; vety 1,008; happi 16,00. Toisen asteen yhtälön ratkaisukaava: ax 2 + bx + c = 0; 1. Kuvassa on esitetty
Valintakoe 2012 / Biokemia Nimi sosiaaliturvatunnus Tehtävissä tarvittavia atomipainoja: hiili 12,01; vety 1,008; happi 16,00. Toisen asteen yhtälön ratkaisukaava: ax 2 + bx + c = 0; 1. Kuvassa on esitetty
Komplementtitutkimukset
 Komplementtitutkimukset Hanna Jarva HUSLAB ja Haartman-instituutti Bakteriologian ja immunologian osasto Komplementti osa luontaista immuunijärjestelmää koostuu yli 30 proteiinista aktivoituu kaskadimaisesti
Komplementtitutkimukset Hanna Jarva HUSLAB ja Haartman-instituutti Bakteriologian ja immunologian osasto Komplementti osa luontaista immuunijärjestelmää koostuu yli 30 proteiinista aktivoituu kaskadimaisesti
KandiakatemiA Kandiklinikka
 Kandiklinikka Pääsykoe 2009 Opiskelijan koe LÄÄKETIETEEN PÄÄSYKOE 2009, OPISKELIJAN KOE Lääketieteen pääsykoe on kuluneina vuosina sisältänyt tehtäviä biologiasta, kemiasta sekä fysiikasta. Pääsykokeen
Kandiklinikka Pääsykoe 2009 Opiskelijan koe LÄÄKETIETEEN PÄÄSYKOE 2009, OPISKELIJAN KOE Lääketieteen pääsykoe on kuluneina vuosina sisältänyt tehtäviä biologiasta, kemiasta sekä fysiikasta. Pääsykokeen
Reseptoripotentiaalista (RP) aktiopotentiaaliin
 aktiopotentiaaliin") Haju- ja makuaisti Reseptoripotentiaalista (RP) aktiopotentiaaliin Reseptoristimulaatio lokaalinen sähköinen ärtyminen (melkein aina depolarisaatio) RP syntymekanismi vaihtelee aistimesta toiseen RP leviää
Haju- ja makuaisti Reseptoripotentiaalista (RP) aktiopotentiaaliin Reseptoristimulaatio lokaalinen sähköinen ärtyminen (melkein aina depolarisaatio) RP syntymekanismi vaihtelee aistimesta toiseen RP leviää
1. Malmista metalliksi
 1. Malmista metalliksi Metallit esiintyvät maaperässä yhdisteinä, mineraaleina Malmiksi sanotaan kiviainesta, joka sisältää jotakin hyödyllistä metallia niin paljon, että sen erottaminen on taloudellisesti
1. Malmista metalliksi Metallit esiintyvät maaperässä yhdisteinä, mineraaleina Malmiksi sanotaan kiviainesta, joka sisältää jotakin hyödyllistä metallia niin paljon, että sen erottaminen on taloudellisesti
Drug targeting to tumors: Principles, pitfalls and (pre-) cilinical progress
 cilinical progress") Drug targeting to tumors: Principles, pitfalls and (pre-) cilinical progress Twan Lammers, Fabian Kiessling, Wim E. Hennik, Gert Storm Journal of Controlled Release 161: 175-187, 2012 Sampo Kurvonen 9.11.2017
Drug targeting to tumors: Principles, pitfalls and (pre-) cilinical progress Twan Lammers, Fabian Kiessling, Wim E. Hennik, Gert Storm Journal of Controlled Release 161: 175-187, 2012 Sampo Kurvonen 9.11.2017
Biopolymeerit. Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä.
 Biopolymeerit Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä. Tärkeimpiä biopolymeerejä ovat hiilihydraatit, proteiinit ja nukleiinihapot. 1 Hiilihydraatit Hiilihydraatit jaetaan mono
Biopolymeerit Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä. Tärkeimpiä biopolymeerejä ovat hiilihydraatit, proteiinit ja nukleiinihapot. 1 Hiilihydraatit Hiilihydraatit jaetaan mono
Perusvuorovaikutukset. Tapio Hansson
 Perusvuorovaikutukset Tapio Hansson Perusvuorovaikutukset Vuorovaikutukset on perinteisesti jaettu neljään: Gravitaatio Sähkömagneettinen vuorovaikutus Heikko vuorovaikutus Vahva vuorovaikutus Sähköheikkoteoria
Perusvuorovaikutukset Tapio Hansson Perusvuorovaikutukset Vuorovaikutukset on perinteisesti jaettu neljään: Gravitaatio Sähkömagneettinen vuorovaikutus Heikko vuorovaikutus Vahva vuorovaikutus Sähköheikkoteoria
Valtimotaudin ABC 2016
 Valtimotaudin ABC 2016 Sisältö Mikä on valtimotauti? Valtimotaudin taustatekijät Valtimon ahtautuminen Valtimotauti kehittyy vähitellen Missä ahtaumia esiintyy? Valtimotauti voi yllättää äkillisesti Diabeteksen
Valtimotaudin ABC 2016 Sisältö Mikä on valtimotauti? Valtimotaudin taustatekijät Valtimon ahtautuminen Valtimotauti kehittyy vähitellen Missä ahtaumia esiintyy? Valtimotauti voi yllättää äkillisesti Diabeteksen