Prolyyli-4-hydroksylaasit
|
|
|
- Jari Halttunen
- 8 vuotta sitten
- Katselukertoja:
Transkriptio

1 Kandidaatintutkielma Prolyyli-4-hydroksylaasit Aleksi Sutinen Oulun yliopisto Biokemian ja molekyylilääketieteen tiedekunta 2016
2 Sisällysluettelo Sisällys I KIRJALLISUUSTUTKIELMA 1. Johdanto Non-hemi rauta(ii)-2-oksoglutara(rankin & Giaccia 2016)atti deoxgenaasien katalysoima reaktio Hypoksia ja normoksia Hypoksian indusoima tekijä Hypoksian indusoiman tekijän rakenne HIF-transkriptiotekijän prolyyli-4-hydroksylaasi HIF-P4H ohjaa HIF1α:n proteasomeihin Kollageenit Kollageenit soluväliaineen osana Kollageenien rakenne Kollageeni prolyyli-4-hydroksylaasi C-P4H:n rakenne ja lokalisoituminen CAT-domeeni osana α-alayksikköä PSB-domeeni osana N-terminaalista domeenia β-alayksikkö-proteiini disulfidi isomeraasi Proliinin hydroksylaation vaikutukset tropokollageeniin Transmembraaninen prolyyli-4-hydroksylaasi P4H-TM:n rakenne ja lokalisoituminen P4H-TM:llä ei ole vielä spesifistä roolia Kirjallisuusluettelo Käytetyt lyhenteet HIF Hypoksia indusoima tekijä HIF-P4H HIF-prolyyli-4-hydroksylaasi HRE Hypoksiaan vastaava elementti EPO Erytropoietiini hormoni
3 VEGF Vaskulaarisen epiteelin kasvutekijä pvhl von Hippel Lindaun proteiini CH-domeenit Kalponiinin kaltaiset domeenit CBP camp:n säätelevään alueeseen liittyvän proteiinin sitoutumisproteiini p300 Adenoviruksen aikasen alueen A1 sitoutumisproteiini C-P4H kollageeni prolyyli-4-hydroksylaasi ARNT Aryyli-nukleaarinen translokaattori ODDD Hapesta riippuvainen hajottava domeeni N/CTAD N-ja C-terminaaliset transaktivaatio-domeenit TRP Tetratrikopeptidi toistuva proteiini FACIT fibrillien kanssa assosioivat ja häiritsevät SCR Nukleaarinen reseptori koaktivaattori II liite: TIEDETTÄ POPULARISOIVA ESITYS Otsikko: Toteutus: Prolyyli-4-hydroksylaasit osana HIF-signalointia ja kollageenien biosynteesiä Posteriesitys
4 1. Johdanto Solun tapahtumia ja reaktioreittejä säädellään tarkoin, jotta solu voi kudoksessa toteuttaa sille kudosspesifistä tehtävää. Solussa käynnistyy signalointitapahtumia, kun solu saa fysikaalisen tai kemiallisen ärsykkeen. Näiden tapahtumien keskiössä ovat entsyymit, joiden biokatalyyttinen kyky mahdollistaa signaalin muuntamisen solun vasteeksi. Vasteella tarkoitetaan muutosta solussa, jonka signaali saa aikaan. Vaste voi ilmetä geenin ekspression käynnistymisenä, solun fyysisenä liikkeenä, tai metabolisena kiihtymisenä. Metabolian kiihtyminen indusoi sellaisia synteesireittejä, jotka vaikuttavat solun kasvuun. Glykolyysi käynnistyy, kun glukoosin määrä plasmassa kasvaa. Jotkin signaalit voivat käynnistää solujen proliferaatiota tai hajoamista. Tässä tutkielmassa perehdytään prolyyli-4-hydroksylaaseihin, joilla on tärkeä rooli molekulaarisen hapen signaloinnissa ja tukikudoksen biosynteesissä. Prolyylihydroksylaasit kuuluvat non-hemi rauta(ii)-2-oksoglutaraattideoksygenaasien superperheeseen, jotka katalysoivat proliini aminohapon hydroksylaatiota. Seitsemänvaiheisen reaktiosarjan seurauksena proliinin pyrrolidiinirenkaan C 4 - hiileen substituoituu hydroksyyliryhmä, jolloin syntyy (2S,4R) hydroksiproliini. Reaktiossa on oltava läsnä orgaaninen koaktivaattori, molekulaarinen O2-happimolekyyli ja askorbaatti (Myllyharju & Koivunen 2013). Hydroksiproliini eristettiin ensimmäisen kerran Gelatiinin hydrolysaateista Emil Fischerin toimesta (Fischer 1902). Proliinitähteiden hydroksylaatio on tärkeä osa P4H kohdeproteiinien post-translationaalista muokkausta. P4H:t toimivat hypoksia-indusoiman tekijän (HIF) ja kollageenien biosynteesin säätelyssä. Eri P4H tyyppejä esiintyy myös muissa eläimissä, kasveissa ja joissakin viruksissa. Kollageenien biosynteesiin osallistuu kollageeni prolyyli-4-hydroksylaasi (C-P4H). Kollageenien perusyksikkönä esiintyy tyypillisesti kolme yhteen liittynyttä α-ketjua, jotka muodostavat helikaalisen rakenteen. C-P4H katalysoi näissä ketjuissa olevien proliini aminohappojen hydroksylaatiota. Hydroksiproliini stabiloi kollageenien rakennetta polaarisen hydroksyyliryhmän vaikutuksesta. Nykyisin tunnettuja ihmisen kollageeni prolyyli-4-hydroksylaasien isomuotoja on kolme, C-P4H (I, II, III). Kollageenien biosynteesin häiriöillä on usein välitön vaikutus luu-, nivel-, rusto- ja epiteelikudosten muutoksiin, mikä tekee C-P4H:n toiminnasta tärkeää (Gorres & Raines 2010). Fysiologista tilaa, jossa solussa ei ole riittävästi happea kutsutaan hypoksiaksi. Vaikeassa hypoksiassa solun alahainen hapen määrä voi johtaa nopeasti akuutteihin kudosvaurioihin. Olosuhteissa jossa hapen määrä on lievästi alentunut, voi pitkään jatkuvana haitata yksilön kehitystä. Tällainen olosuhde 1
5 vallitsee esim. sikiökauden aikana jossa hapen saanti on rajoittunutta äidin istukan kautta. Elimistössä hypoksiavaste käynnistää hypoksia indusoima tekijän (HIF) aktivoitumisen. HIF:it ovat joukko transkriptiotekijöitä, jotka säätelevät noin sataa kohdegeeniä. Näillä geeneillä on kiihdyttävä vaikutus solun metaboliaan ja kasvuun. Näitä transkriptiotekijöitä tunnetaan kolmea eri isomuotoa, HIF (1,2,3). HIF transkriptiotekijöiden liiallinen aktiivisuus johtaa kudosten rajoittamattomaan kasvuun, joka voi edesauttaa syöpäsolujen muodostumista (Semenza 2012). Hypoksialla ja HIF-ohjautuvien geenien yliekspressioilla on yhteys vaikeiden tuumorien kehittymiseen ja etäpesäkkeiden muodostumiseen (Rankin & Giaccia 2016). Normoksia on fysiologinen tila, jossa happea on solussa riittävästi. Tällöin solu pyrkii rajoittamaan HIF-ohjautuvien geenien yliekspressioita. HIF-signaalin vaikutusta säätelee solussa HIF-prolyyli-4-hydroksylaasit. Ne hydroksyloivat HIF:n kahdessa transaktivaatiodomeenissa olevia proliinitähteitä, jolloin HIF:n sitoutuminen kohdegeenin säätelyalueelle estyy. 1.1 Non-hemi rauta(ii)-2-oksoglutara(rankin & Giaccia 2016)atti deoxgenaasien katalysoima reaktio Prolyyli-4-hydroksylaasien katalysoima reaktio tapahtuu non-hemi-rauta(ii)-dioksigenaasien reaktiomekanismin mukaisesti. Seitsemänvaiheinen reaktiosarja (kuva.1) on monimutkainen ja se voidaan jakaa kahteen osaan. Ensin muodostuu reaktiivinen rauta(iv)-superoksidi, jota seuraa substraatin hydroksylaatio ja koentsyymin dekarboksylaatio (Costas et al. 2004). Reaktiomekanismin yksityiskohtaisen mekanismin esittivät ensimmäisen kerran Hanauske-Abel ja Günzler vuonna (1982). Reaktiota katalysoivan entsyymin katalyyttiset aminohapot ovat aspartaatti tai glutamaatti, sekä kaksi proliinia. Nämä kolme aminohappoa muodostavat kolmikon samalle puolelle metallikompleksia. Toisen puolen muodostavat koaktivaattori ja molekulaarinen happi. Ensimmäisessä vaiheessa muodostuu reaktiivinen ferryylikompleksi, jonka keskusatomina on rauta(iv)-kationi. Reaktion alkaessa keskusatomin hapetusluku on vielä kaksi. 2-oksoglutaraatti (ts. α-ketoglutaraatti) toimii reaktion koentsyyminä. Se muodostaa ligandin rauta(ii)-kationiin C 1 -hiilen karboksylaattiryhmän hapella ja C 2 -hiilen karbonyyliryhmän hapella. Askorbaatti pelkistää Fe(III):sta Fe(II):ksi. Tämä on reaktion kannalta oleellista, koska rauta(iii):n redox-potentiaali ei ole reaktion kannalta riittävä (de Jong et al. 1982). Happimolekyylin sitouduttua aktiiviseen kohtaan, sen toiselle happiatomille syntyy negatiivinen varaus, jolla se hyökkää 2-oksoglutaraatin karboksyyliryhmän hiileen. Tällöin muodostuu Fe(IV)- 2
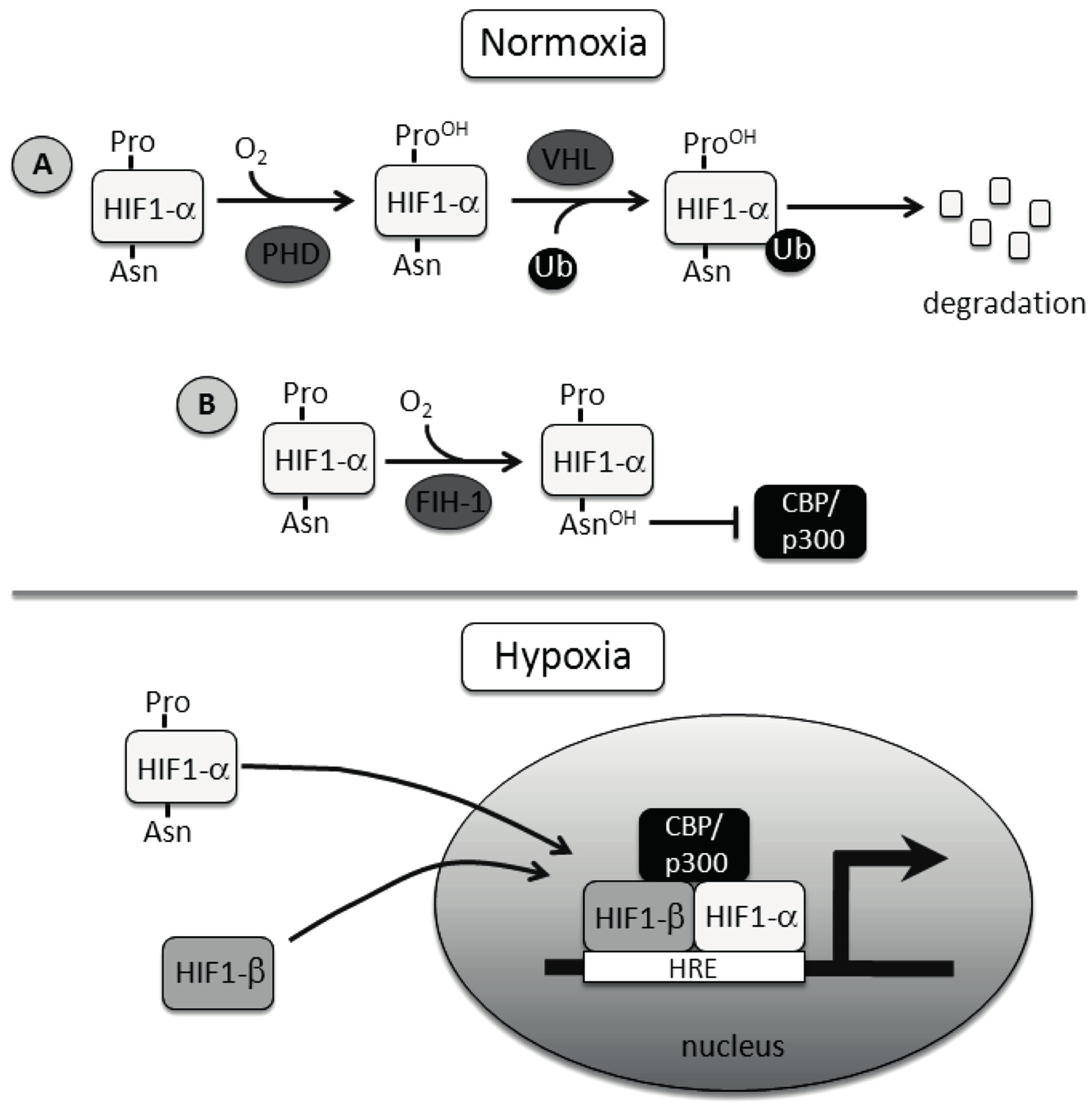
6 peroksihemiketaali-kompleksi, jossa keskusatomi rauta on sitoutunut viiteen substituenttiin. 2- oksoglutaraatin karboksyyliryhmä dekarboksyloituu muodostaen hiilidioksidia. Kompleksissa olevan ketoryhmän happiatomin affiniteetin ansiosta proliinin pyrrolidiinirengas radikalisoituu. Tällöin pyrrolidiinirenkaan C 4 -hiilestä irtoaa ensin protoni, joka korvautuu ketoryhmän happiatomilla. Pyrrolidiinirenkaan C 4 -hiileen substitioutuu hydroksyyliryhmä ja koaktivaattori dekarboksyloituu sukkinaattina (Gorres et al. 2008, Hoffart et al. 2006, Krebs et al. 2007) Kuva.1 Non-hemi-Fe(II)-2-oksoglutaraatti dioksigenaasien katalyysin reaktiomekanismi. Rauta kelatoituu kahden histidiinin ja yhden aspartaatin kanssa samalle puolelle. Toinen puoli ligandista muodostuu 2-oksoglutaraatin karbonyyliryhmän ja karboksylaatin happiatomeista. Substraatin lähestyessä katalyyttistä aluetta molekulaarinen happi liittyy kompleksiin ja hyökkää 2-oksoglutaraatin karbonyylihiileen, jolloin tapahtuu dekarboksylaatio ja syntyy hiilidioksidia. Muodostuu reaktiivinen ferryyli-fe(iv)-metallikompleksi, jonka seurauksena muodostuu prolyyliradikaali. Tämän jälkeen happi liittyy pyrrolidiinirenkaaseen ja sukkinaatti dekarboksyloituu. Lopputuotteena syntyy hydroksiproliini, sukkinaatti ja vesimolekyyli. (Kuva lähteestä: (Tarhonskaya et al. 2014). Kuvan käyttöön on anottu lupa kustantajalta. Teksti lähteistä: Costas et al. 2004, Hoffart et al. 2006, McDonough et al. 2006, Gorres et al. 2008) 2. Hypoksia ja normoksia 2.1 Hypoksian indusoima tekijä EPO-hormonin säätelyyn liittyviä havaintoja voidaan pitää ratkaisevana hypoksia-indusoiman tekijän löytymisen kannalta. EPO-geenin löytyminen ja sen jatkotutkimukset johtivat ensimmäiseen hapen avulla ohjautuvan transkriptiosäätelyalueen HRE:n (Hypoxia responsive element) löytymiseen (Beck et al. 1991, Lin et al. 1985). HRE-alueeseen liittyvien transkriptiotekijöiden selvittäminen johti HIF:n löytymiseen. Nykyisin tunnetaan kolmea eri HIF-isomuotoa, joista HIF1α karakterisoitiin 3

7 ensimmäisenä (Wang et al. 1995). HIF:t voivat indusoida angiogeneesin kannalta tärkeiden prekursorien kuten EPO-hormonin ja VEGF-hormonin ekspression käynnistymisen (Kuva.2). Muita kasvua stimuloivia tekijöitä ovat mm. IGF- ja TGF-hormonien tuotannon lisääminen. Solun metaboliaan HIF:t voivat vaikuttaa lisäämällä mm. GLUT1-transportterien määrää. Hapen kuljetusta verisuonissa voidaan tehostaa, kun HIF:t indusoivat vasomotorisia kohdegeenien kuten inos:n tuottoa. Hapen merkitys solun homeostaasiin on ollut tiedossa pitkään, mutta sen mekanismeja ollaan alettu ymmärtämään molekulaarisella tasolla vasta viime vuosien aikana (Rankin & Giaccia 2008, Myllyharju & Koivunen 2013). Kaksi muuta HIF-isomuotoa ovat HIF2α ja HIF3α (Gu et al. 1998, Wenger & Gassman 1997) Kaikki HIF:t muodostavat αβ-heterodimeerejä, joissa β-alayksikkö on kaikissa isomuodoissa identtinen. Kuva.2 HIF-signaloinnin pääperiaatteet. Happi toimii signaalivälitteisessä solun säätelyssä Hypoksian indusoiman tekijän avulla. Hypoksiassa sytoplasmassa olevan HIF1α:n määrää säätelevä HIF-P4H inhiboituu. Tämä johtaa HIF1α:n kulkeutumisen tumaan, jossa se dimerisoituu HRE-säätelyalueeseen HIF1β:n ja p300/cbp:n kanssa. Nämä transkriptiotekijät indusoivat noin sadan eri geenin transkriptiota, jotka vaikuttavat kudosten metaboliaan kiihdyttävästi. Normoksisessa tilassa happi aktivoi yhdessä Fe 2+ :n 2-oksoglutaraatin ja askorbaatin kanssa HIF-P4H-entsyymin joka hydroksyloi HIF1α:n kahta proliini aminohappoa. Hydroksylaation seurauksena pvhl-kompleksi ohjaa HIF1α:n proteosomaaliseen hajotukseen. (Kuva lähteestä(maes et al. 2012). Kuvaan on saatu lupa kustantajalta. Teksti lähteestä: (Semenza 2001) 4
8 Laajan kohdgeenijoukon ansiosta hypoksia kytkeytyneellä säätelymekanismilla on suuri merkitys solun homeostaasiin, solunjakautumiseen ja apoptoosiin (Semenza 2001). Laajan geenijoukon vuoksi HIF signalointi voi johtaa usein onkologisten prosessien käynnistymiseen, joiden tutkiminen on avannut mahdollisuuksia tiettyjen syöpien kliinisten hoitomenetelmien kehittämiseen (Kung et al. 2000). 2.2 Hypoksian indusoiman tekijän rakenne Hypoksiassa solu käynnistää sensorijärjestelmän, jonka keskeisessä roolissa on HIF1α- ja HIF1βtranskriptiotekijöiden aktivoituminen, jota kontrolloidaan 2-oxoglutaraatti riippuvaisten dioxygenaasientsyymien hydroksylaatio reaktion avulla (Bishop & Ratcliffe 2014). Pääasiallisesti α- alayksikön stabiliteetti on olennainen osa reaktioreitin säätelytapahtumaa. HIF:it kuuluvat PASproteiinien superperheeseen, joissa esiintyy tavallinen loop-helix-loop PAS laskostyyppi (Wang et al. 1995). DNA:han sitoutuva osa on happisensorina toimiva α-alayksikkö ja geenin säätelyalueeseen kiinnittyvä β-alayksikkö. Dimerisoituminen kohdentuu HIF1α:n N-terminaaliseen osaan (aminohapot 1-390). DNA:han sitoutuva osa on myös kyseisessä sekvenssissä (Jiang et al. 1996). HIF1:n aktivoituminen saa aikaan satojen hypoksia-kytkettyjen geenien ekspression, joka ohjaa mm. glykolyysiä, angiogeneesiä, erytropoietiisiä ja vaskulariteettiä. PAS-proteiinit koostuvat kolmesta alayksiköstä, PER- (period circadian protein), AHR- (aryl-hydrocarbon receptor) / ARNT (arylhydrocarbon nuclear translocator) ja SIM (single-minded protein) proteiineista. HIF1α:ssa esiintyvä β-domeenilla on ARNT aktiivisuus (Hoffman et al. 1991). Hypoksisessa tilassa HIF1α stabiloituu ja muodostaa heterodimeerin β-alayksikön kanssa, joka toimii tässä tilanteessa ARNT:n kaltaisesti (aryl-hydrocarbon nuclear translocator) (Hoffman et al. 1991, Wang et al. 1995). HIF1 aktivaatio vaatii lisäksi useiden avustavien transkriptiofaktoreiden liittymistä HRE-säätelyalueelle, joita ovat p300, CBP, tai SRC (Kallio et al. 1998, Kung et al. 2000). Näillä proteiineilla on histoni trans-asetyyli-transferaasi aktiivisuutta. HIF1-dimeerin ja avustavien transkriptiotekijöiden interaktiot tapahtuvat HIF-dimeerin C- terminaalisen transaktivaatio domeenin (CTAD), N-terminaalisen transaktivaatio domeenin (NTAD) ja p300/cbp proteiineissa esiintyvien CH1, CH2 ja CH3 domeenien välillä. Transaktivaatiodomeenit sijaitsevat vierekkäin HIF1α:n rakenteessa järjestyksessä, jossa N-terminaalinen transaktivaatio-domeeni tulee ennen C-terminaalista. Niitä yhdistää N-terminaalisesta ulottuva looprakenne. Muut CTAD:n rakenteelliset osat ovat kaksi helixiä ja niiden väliin muodostuva looprakenne. CH-domeenit sisältävät kysteiini/histidiini motiivin joiden avulla p300/cbp liittyy HIF- 5
9 dimeerin C-terminaaliseen transaktivaatio-domeeniin. CTAD:n ja CH1:n kompleksi on orientoitunut kolmion muotoon katalyyttisessä keskuksessa. (McDonough et al. 2006) CH1 ei kuitenkaan muodosta sidosta CTAD:n rikkisillan avulla vaan sidos muodostuu lähinnä hydrofobisten aminohappojen vuorovaikutusten avulla. Lisäksi on havaittu, että kysteinit kelatoivat Zn 2+ -kationin inhibitorisesti hydrofobisen taskun sisään, joka välttämätöntä kompleksin muodostukseen. p300/cbp ja HIF-dimeerin sitoutuminen CTAD sijoittuu aminohappojen välille (Freedman et al. 2002). Normoksiassa HIF1:n dimerisoituminen estyy hapen feedback-inhibition seurauksena. Tällöin HIF1α ei pysy stabiilina, eikä se voi siten muodostaa kompleksia HIF1β:n (ARNT) kanssa. Epästabiili HIF1α jää sytoplasmaan, jossa se hydroksyloidaan HIF-P4H:n katalysoimassa reaktiossa (Semenza 2001). pvhl (Von Hippel-Lindau protein) on Von Hippel-Lindaun kasvunrajoittajageenin tuote. Se ei yleensä esiinny solussa monomeerinä, vaan kompleksissa elongiini C:n ja elongiini B:n kanssa (Kibel et al. 1995). Tällä kompleksilla on E3-ubikitiini-ligaasi aktiivisuutta. pvhl on αβ-heterodimeerinen proteiini. Sen α-domeeni rakentuu kolmesta pienestä α-kierteestä ja β-domeeni useammasta β- laskoksesta. Domeeneja yhdistää kaksi silmukkaa. Elongiini C on suurimmaksi osaksi kohdakkain pvhl:n α-domeenin kanssa (Stebbins et al. 1999). HIF1α:ssa oleva ODDD (Oxygen dependent degradation domain) säätelymotiivi liittyy pvhl:n β-domeeniin (Ohh et al. 2000). Katalyyttiset proliinit, joihin hydroksylaatio kohdistuu ovat Pro 402 ja Pro 564 (Epstein et al. 2001). Proliinien läheisyydessä olevat aminohapot ovat erilaiset kollageenien X-Y-Gly-jaksoihin nähden. HIF- P4H:den hydroksylaatio kohdistuu Leu-X-X-Leu-Ala-Pro-X-motiiveihin (Huang et al. 2002). Reaktion mekanismi noudattaa aikaisemmin esiteltyä 2-oksoglutaraattidehydrogenaasien tyypillistä reaktiota, jossa on oltava kofaktorina rautaa(fe 2+ ) ja askorbaattia. Lisäksi raudan sitomiseen tarvitaan 2-oksoglutaraattia ja happea (Jaakkola et al. 2001). HIFα:n isomuodot ekspressoituvat eri kudoksissa ja niiden geenit ovat konservoituneita siten, että ne muokataan vaihtoehtoisen silmukoinnin avulla (Epstein et al. 2001). HIF1α ja HIF2α ovat EPASproteiineja (Endothelial PAS) (Tian et al. 1997) ja HIF3α ovat IPAS (Inhibitory PAS) (Makino et al. 2001)- tai NEPAS-proteiineja (Neonatal and embryonic PAS) (Yamashita et al. 2008) Näiden eri PAS proteiinien ekspression esiintyvyydet eri kudoksissa eroavat hiukan, mutta pääasiallisesti HIF:ien ekspressio on suurinta aivokuoren-, haiman-, istukan- ja luuytimen soluissa (Shan et al. 2012). 6
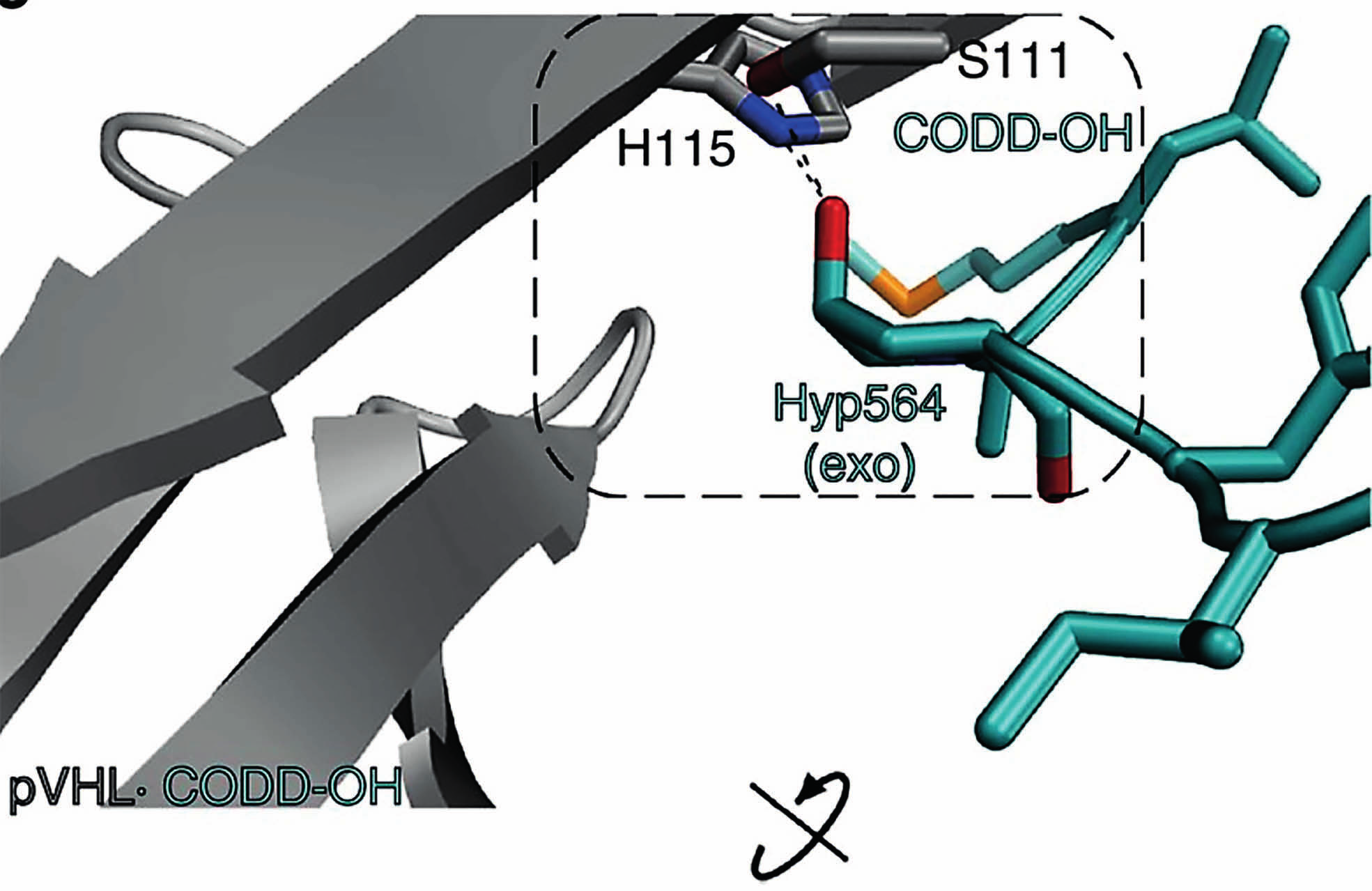
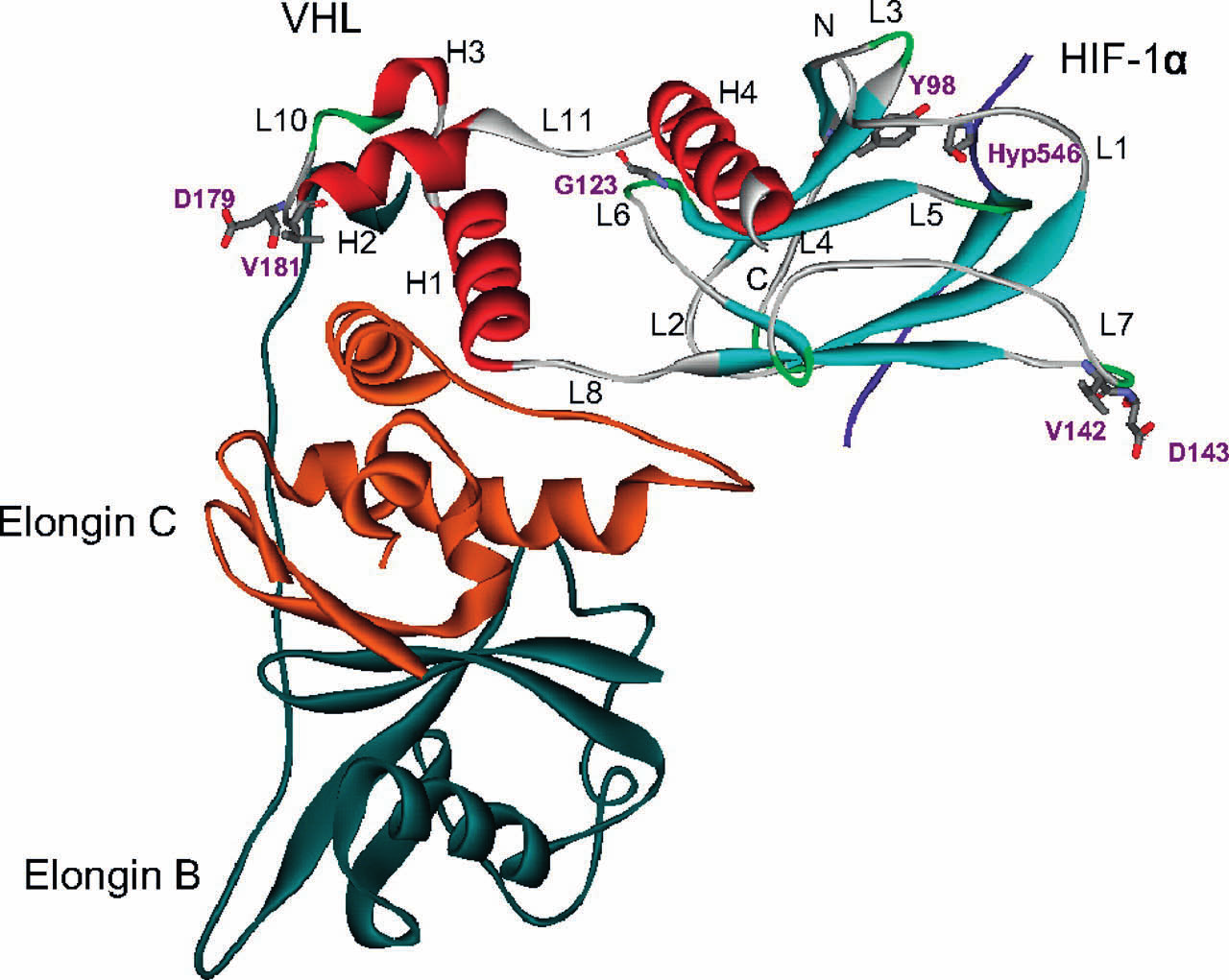
10 Erityisesti sikiön kehityksen aikana sikiön solukko on altistuneena hypoksialle, josta johtuen varsinkin HIF1α ja HIF2α ekspressiot ovat suuria istukan solukossa. HIF3α on allosteerinen HIF2α:n ja HIF1α:n säätelijä. Se muodostaa myös heterodimeerin ARNT-yksikön kanssa ja liittyy HREsäätelyalueeseen, mutta sen affiniteetti on huomattavasti heikompi kuin HIF1α:n tai HIF2α:n Heikompi sitoutuminen säätelytekijään on selitettävissä sillä, että HIF3α: geenissä on C- terminaalisen trans-aktivaatiodomeenin sijasta, vain N-terminaalinen trans-aktivaatio domeeni. Se siis kilpailee ARNT kompleksinmuodostuksesta HIF1α: ja HIF2α:n kanssa. Tämän säätelymekanismin häiriöillä on havaittu olevan yhteyttä neonataalien elinvaurioihin tai kasvun häiriöihin (Compernolle et al. 2002). Toinen HIF3α variantti NEPAS toimii erityisesti vastasyntyneen keuhkojen epiteelisoluissa, toimien HIF1α:n ja HIF2α:n negatiivisena säätelytekijänä. Havainnot HIF3α:n ekspression indusoitumisesta vastasyntyneillä, voidaan selittää vastasyntyneen luontaisen kardiorespitoriaalisen hengityksen käynnistymisellä. Sikiökauden aikana äidin istukan kautta saatu hapen määrä on usein rajoittunutta sikiön kehitykselle, jolloin HIF1α ja HIF2α ovat aktiivisia. Tämä on välttämätöntä mm. sikiön vaskulariteetin ja angiogeneesin kannalta (Iyer et al. 1998, Ryan et al. 1998). Synnytyksen jälkeen hapen määrä kasvaa dramaattisesti, jolloin HIF-ohjatuvien geenien indusointi ei ole enää tarpeellista. HIF1α:n ODDD:ssä sijaitsevien proliinitähteiden lisäksi samankaltainen hydroksylointi voi tapahtua toiseen konservoituneeseen aminohappoon CTAD:ssa, joka on Asparagiini. Tämän posttranslationaalisen modifikaation tarkoitusperä on sama kuin proliinien hydroksylaatiolla. Näin ollen se toimii HIF1α säätelyssä negatiivisesti. Yleisesti asparagiini-hydrokslylaasia kutsutaan FIH:ksi (Factor inhibiting HIF). p300/cbp-kompleksi, liittyy CTAD:iin, mikäli Asp 803 :ta ei ole hydroksyloitu. Entsyymi, joka katalysoi tämän reaktion on aspartyyli-β-hydroxylaasi, joka kuuluu myös non-hemi rauta(ii)-2-oxoglutaraatti riippuvaisiin dioksygenaaseihin (Wang et al. 1991). Tämä mahdollistaa CTAD:n aktivoitumisen ja HIF1αβ-dimeerin muodostumisen. HIF-indusoitu geeniekspressio voidaan kiteyttää kahteen vaiheeseen. Jotta mekanismi voi käynnistyä, kaksi seikkaa on tapahduttava: HIF1α:n Pro 402 ja Pro 564 hydroksylaation inhibointi ja Asp 803 hydroksylaation inhibointi. Nämä seikat mahdollistavat sen, että pvhl-kompleksi ei voi sitoutua HIF1α:n proliineihin, jolloin p300/cbp pääsee liittymään CTAD:iin (Lando et al. 2002). 7
11 3. HIF-transkriptiotekijän prolyyli-4-hydroksylaasi 3.1 HIF-P4H ohjaa HIF1α:n proteasomeihin Normoksisessa tilassa HIF1α:a hajotetaan tehokkaasti proteasomeissa ubikitiniini-kytketyllä systeemillä, joka estää HIF1α:n pääsyn tumaan (Huang et al. 1998). Ennen kuin tiedettiin mekanismista, jolla voitiin selittää HIF1α:n hajottamista proteasomeissa, tiedettiin että metallikationit kuten Co 2+, Ni 2+, Mn 2+ ja rautaa kelatoivat yhdisteet, kuten EDTA kykenivät aktivoimaan HIF1α:a liittymällä CTAD:n ja inhiboimaan HIF-P4H:ta (Ivan et al. 2002). (Myllyharju & Koivunen 2013)(Myllyharju & Koivunen 2013)(Rankin & Giaccia 2008)Proteiinien hapettumista koskeva reaktiomekanismi, jossa metallikationit toimivat katalyyttinä tunnettiin (Hanauske-Abel & Günzler 1982, Levine et al. 1981, Levine 1983).Metallikationien inhiboiva vaikutus antoi viitteitä sille, että HIF1α:n säätelyyn osallistuu sellainen entsyymi, jonka katalyysissä HIF1α modifioidaan oksidatiivisesti. Entsyymikatalyysin, jossa ODDD-domeenin P 402 ja P 564 hydroksyloidaan, oletettiin olevan riippuvainen hapesta ja raudasta. Tämä pystyttiin osoittamaan kokeellisesti (Ivan et al. 2001). HIF-P4H:t ovat Fe +2-2-oxoglutaraatti riippuvaisia dioksygenaaseja. Sen katalyysi perustuu samaan mekanismiin kuin C-P4H:n α2β2 tetrameerin (Kivirikko & Pihlaniemi 1998) protokollageenin hydroksylointi, mutta substraattia ympäröivien aminohappojen orientaatio proliinien ympärillä on erilainen. Lisäksi C-P4H:n katalyysi tapahtuu ER:ssa ja HIF-P4H:n sytosolissa, eikä sen toiminta esty hypoksisessa tilassa (Takahashi et al. 2000). C-P4H ei kuitenkaan voi hydrolysoida HIF1α:n proliineja. Kaikki P4H:n aminohapot, jotka sitovat Fe 2+ -ionia ovat konservoituneita. Tämän perusteella pystyttiin osoittamaan kokeellisesti HIF-P4H:n aktiivisuuden moninkertaistuvan askorbaatin ja FeSO 4+ :n vaikutuksesta (Bruick & McKnight. 2001). Samalla myös pystyttiin osoittamaan, että CoCl2 inhiboi hydroksylaatiota, sillä se kilpailee Fe 2+ :n sitoutumisesta HIF-P4H:n katalyyttiseen motiiviin. Homologien perusteella ja käyttämällä bioinformatiivisia menetelmiä C.eleganssin ja D.melanogasterin geenisekvensseistä, Egl9 geenistä löydettiin ihmisen HIF-P4H:ta. Nykyään tunnetaan kolmea HIF-P4H:n isomuotoa. Ne esiintyvät kirjallisuudesta riippuen eri nimillä, joita ovat HIF-P4H:den (1,2,3) lisäksi, mm. PHD:t (1,2,3) ja EGLN:t (1,2,3) (Bruick & McKnight. 2001, Epstein et al. 2001, Ivan et al. 2002). HIF-P4H:n ja C-P4H:n katalyyttisen domeenin laskostyyppi on samankaltainen. Se koostuu kahdeksasta β-laskoksesta, jotka asettuvat päällekkäin (double stranded β-helix, jelly-roll-fold). 8
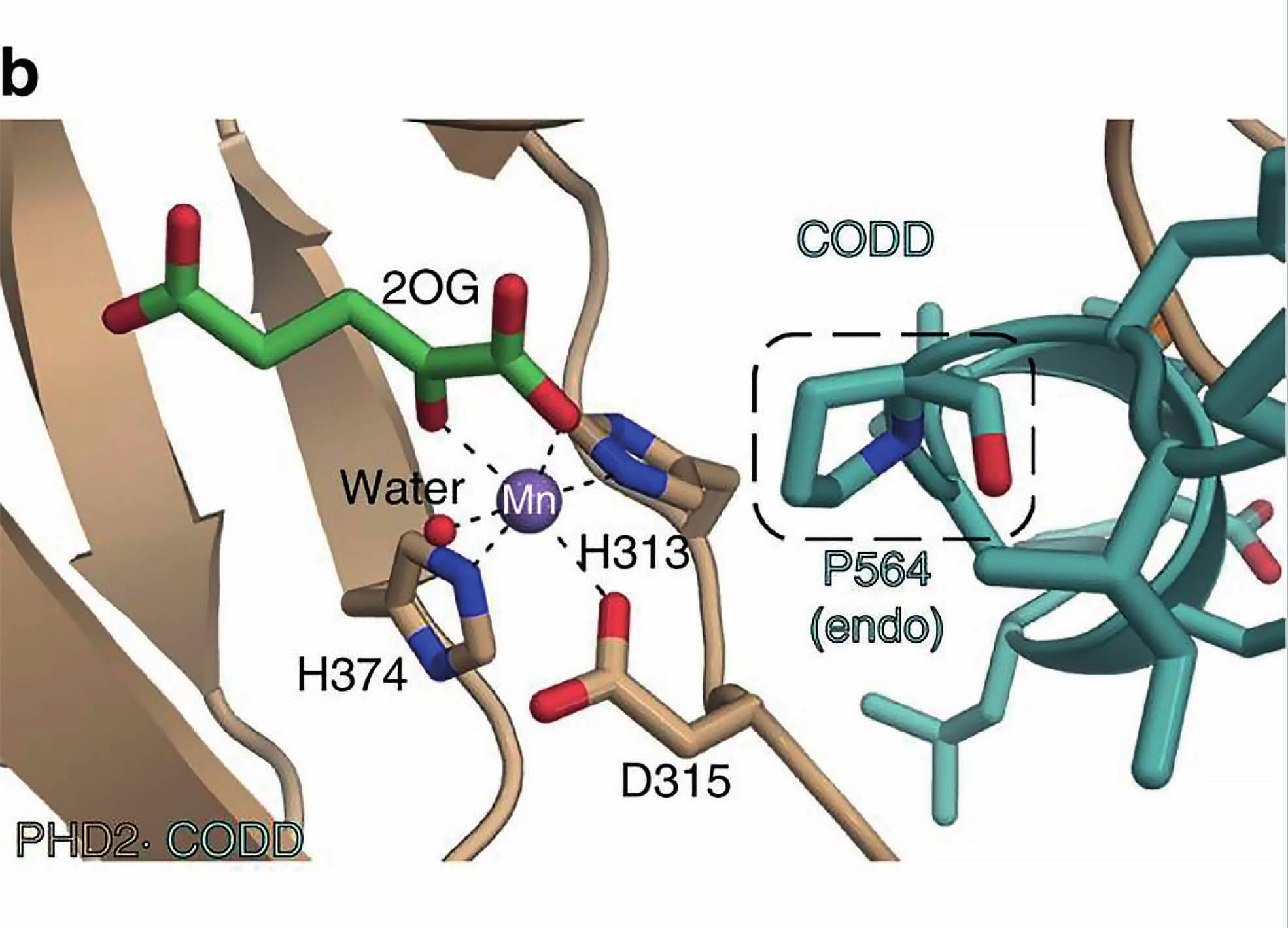
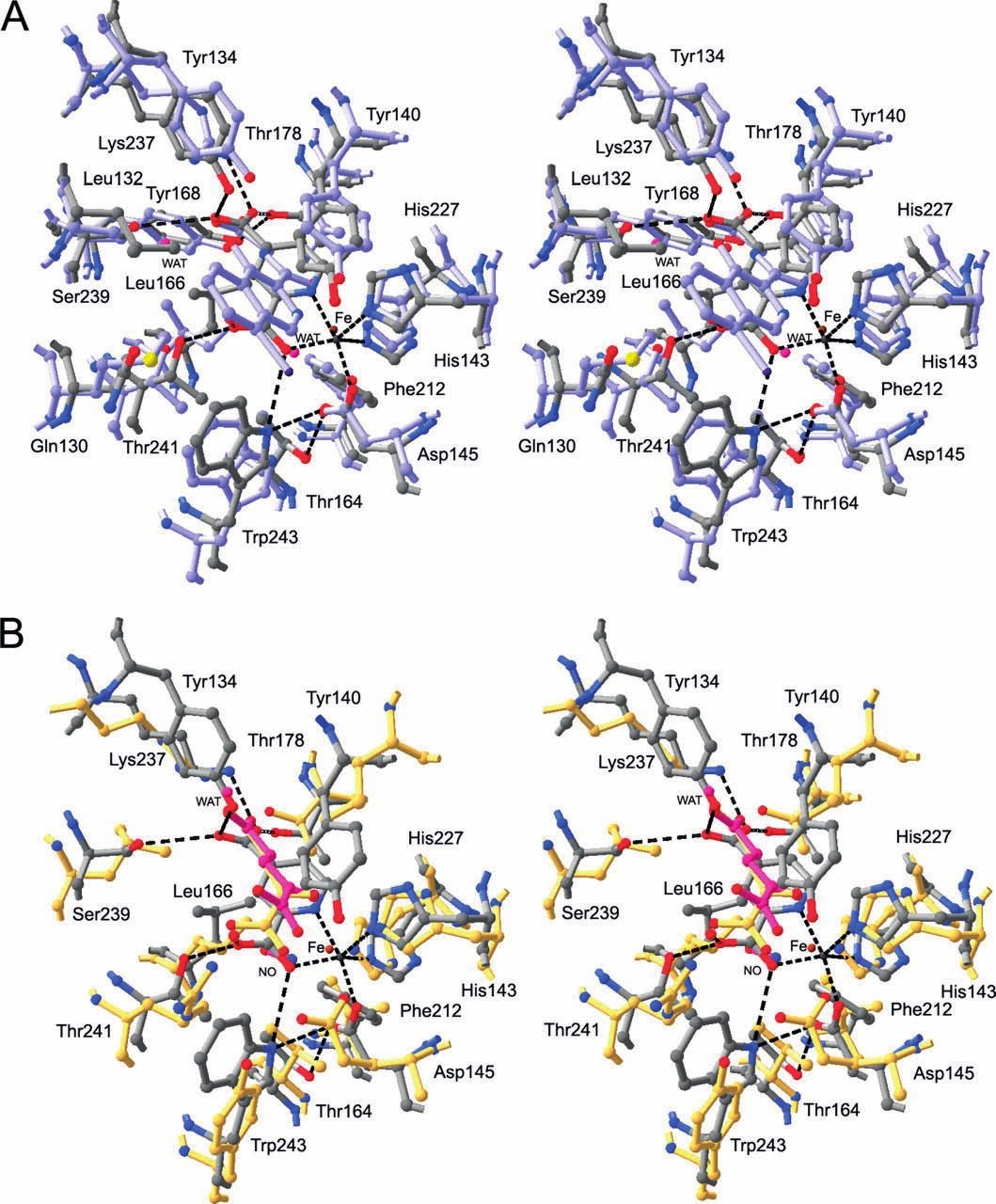
12 Katalyysin kannalta tärkeät aspartaatti ja histidiinit tulevat toisen β-laskoksen jälkeen. Kaikkia β- laskoksia stabiloi kolme ympäröivää α-kierrettä. Entsyymi esiintyy solussa monomeerinä. Sen katalyyttisen yksikön rakenenteessa rauta(ii)-kationi on kompleksoitunut oktaedriesesti His 313 :n, Asp 315 :n ja His 374 :n, 2-oksoglutaraatin ja happimolekyylin kanssa (McDonough et al. 2006). 4. Kollageenit 4.1 Kollageenit soluväliaineen osana Solu tarvitsee monimutkaisten säätelyreittien lisäksi muodon ja liikkeen. Kudoksen täytyy kasvaa ja toisaalta antaa tilaa ja joustaa. soluväliaine rakentuu epiteeli-, tai non-epiteelikudoksesta ja se rajoittuu laminaan. Laminaa ylläpitää sen kanssa vuorovaikuttavat solut. Kaikilla eliöillä on tarve uusiutua, siten soluväliaineen rakenneproteiineissa esiintyy eläinkunnassa paljon homologisia rakenteita. Sen pääkomponentteja ovat joukko suurikokoisia proteiineja, jotka voidaan jakaa neljään luokkaan, tyypin IV kollageenit, laminiinit, perlekaanit ja nidokiinit. Näistä kollageenit esiintyvät runsaimpina kaikissa eliöissä. Ihmisen koko proteiinimassasta 30% on kollageenia. Kollageenien biosynteesi tapahtuu pääosin fibroplasteissa. Kaikkiaan kollageeneja tunnetaan 27 erilaista tyyppiä, joista tyypin I kollageeni on kaikista yleisin selkärankaisilla tavattu kollageeni. Tyypin I kollageeni muodostuu kahdesta α-ketjusta, jotka kummatkin koostuvat n. 1000:sta aminohaposta ja ovat nm pitkiä. Kollageenit syntetisoidaan monomeerisena prokollageeneina, jotka rakentuvat fibrillisäikeiksi. Posttranslationaalisen muokkaamisen jälkeen kollageenia kutsutaan tropokollageeniksi. Polaarisuuden tuoman lujuuden ansiosta kollageeni on luu-, nivel- ja sidekudoksen pääkomponentti. Tyypin IV kollageenit esiintyvät laminan peruselementtinä. Muita kollageenityyppejä, joita esiintyy eri kudostyypeistä riippuen ovat: säikeitä muodostavat-(fibrillaariset), FACIT-(fibril associated collagens with interrupted triple helix), heksagonaaliset-, ankkuroivat-, transmembraaniset kollageenit ja multipleksiinit. Lisäksi on olemassa joukko kollageeneja, joiden toimintaa ei toistaiseksi tunneta vielä hyvin (Shaw & Olsen 1991). Polaarisuuden tuoman lujuuden ansiosta kollageeni on luu-, nivel ja sidekudoksen pääkomponentti. Noin 80-90%:a ihmiskehon kollageeneista on fibrillaarisia (tyypit I, II ja III). Fibrillaaristen kollageenien kanssa interaktoivat FACIT-kollageenit liittävät näitä soluväliaineen muihin komponentteihin ja epiteelikudoksiin. Transmembraanisten kollageenien tehtävänä on muodostaa liitoksia (neksus) solujen välille. Ne nopeuttavat solujen välistä viestintää, joista desmosomit ovat yleisin esimerkki. Kollageeneilla voi olla myös lymfaattisia ja proteoglykaanisia ominaisuuksia (Shaw & Olsen 1991). 9
13 4.2 Kollageenien rakenne Kollageenit ovat trimeereistä koostuvia polymeerejä, jotka jakaantuvat α-ketjuiksi. Kudostyypistä riippuen α-ketjut voivat olla homotrimeerejä tai heterotrimeerejä. Ketjut kiertyvät toistensa kanssa kolminkertaiseksi kierteeksi muodostaen fibrillaarisia tai verkkomaisia tertiääri-rakenteita. Prokollageenien hydroksylaatio lisää ketjujen polaarisuutta, jonka ansiosta tropokollageeni saavuttaa sille ominaisen lujuuden. kollageenit ovat luu-, nivel ja sidekudoksen pääkomponentteja (Heino & Vuento 2009). Kollageenin primäärinen helix-rakenne perustuu X-Y-Gly-jaksoihin (Bowes & Kenten 1948). Toistuvan yksikön ainoana vaatimuksena joka kolmannen aminohapon tulee olla glysiini. Kaksi muuta aminohappoa voivat olla mitä tahansa aminohappoja, mutta usein niitten paikalla ovat proliini ja hydroksiproliini. Glysiini on tärkeä osa kollageenin rakennetta, koska sen pieni koko ja vähäinen steerinen este sallivat α-ketjujen tiiviin pakkautumisen. Kaksi muuta aminohappoa ovat polaarisia glysiiniin nähden, joiden rooli on stabiloida α-ketjua. Post-translationaalisen modifikaation seurauksena proliinista muodostuu hydroksiproliini, kun proliinin pyrrolidiinirenkaaseen liittyy hydroksyyliryhmä. Happi vetää negatiivisen osittaisvarauksen avulla elektroneja kohti akselin keskiosaa, stabiloiden α-ketjun rakennetta. Ketjut liittyvät toisiinsa vetysidoksin (Lodish et al. 2007). Kollageenityyppien erot voidaan jakaa kolmeen eri karakteristiseen tekijään. Suurin ero kollageeneillä on niiden kolmoiskierteisten α-kierteiden määrässä ja pituudessa. Yksittäinen kollageeniyksikkö ei ole koko matkaltaan kolmoiskierteinen. Välissä voi olla sivuavia, tai niiden väliin sijoittuvia segmenttejä, jotka eivät ole kierteisiä. Kiertymättömien ketjujen ansioista kollageenit saavat elastisen ominaisuuden. Kolmas ero kollageenien välillä riippuu kovalenttisesta luonteesta ketjujen välillä. Proliinin hydroksylaation lisäksi α-ketjua voidaan glykosyloida, hapettaa, tai ketjut voivat mennä ristiin toisten ketjujen kanssa. Ketjuun voi liittyä myös globulaarisia yksiköitä. Kollageenit ovat rakenteeltaan hyvin primitiivisiä, mutta suuria proteiineja. Niillä on erittäin tärkeä rooli soluväliaineen toiminnassa ja niiden virheellisellä laskostumisella on usein aina yhteys tuki-ja liikuntaelimistön sairauksiin. (Lodish et al. 2007). 5. Kollageeni prolyyli-4-hydroksylaasi 5.1 C-P4H:n rakenne ja lokalisoituminen Toinen tärkeä prolyyli-4-hydroxylaaseihin kuuluva entsyymi HIF-P4H:n ohella on kollageenin X-Y- Gly-jaksojen proliineja ja muita kollageenin tyyppisiä peptidejä hydroksyloiva kollageeni-prolyyli- 10
14 4-hydroksylaasi (C-P4H) (Kivirikko 1989). Sen katalyysin suorittava α-alayksikkö kuuluu myös nonhemi rauta(ii)-2-oksoglutaraatti dioksygenaaseihin. Sen konformaatiota ylläpitää tioreduktaaseihin kuuluva proteiini disulfidi isomeraasi, joka muodostaa β-alayksikön. Näin ollen kaikki selkärankaisten C-P4H:t ovat α2β2 heterotetrameereja. Ihmisen C-P4H α(i)-alayksikkö koostuu 517:ta aminohaposta ja 17 aminohapon pituisesta signaalijaksosta, kun taas α(ii)-alayksikkö koostuu 514:ta aminohaposta ja 21:n aminohapon pituisesta signaalijaksosta (Annunen et al. 1997, Helaakoski et al. 1989). C-P4H:n toiminnallisuudessa ei ole yhteyttä HIF-ohjautuvien geenien säätelyyn ja sen geenin identtisyys HIF-P4H:den kanssa on vähäinen. Täysin eri tehtävästä huolimatta entsyymeillä on konservoituneita aminohappoja ja samankaltainen katalyyttinen mekanismi. Niiden substraatit ovat erilaisia, joten niiden katalyyttisen domeenin aminohappojen orientaatio on erilainen. Toisin kuin HIF-P4H:t, C-P4H:t lokalisoituvat ER:n luumeniin. C-P4H osallistuu prokollageenin posttranslationaalisessa muokkaukseen (Kivirikko & Prockop 1967). Nämä seikat ovat auttaneet paljon HIFα (1,2,3):n ja HIF-P4H(1,2,3):n tutkimuksissa. Ihmisen C-P4H:n ohella entsyymiä on löydetty ja tutkittu, myös muista selkärankaisista, kuten C.eleganssista ja D.melanogasterista. Eläimien lisäksi C-P4H:ta on löydetty kasveista mm. Arabidopsis thalianasta (Hieta & Myllyharju 2002) ja Chlamydomonas reinhardtiista (Koski et al. 2009). Joittenkin viruksien, kuten Paramecium bursaria Chlorella Virus-1:n katalyyttinen mekanismin osoittava kiderakenne on pystytty ratkaisemaan kristallografian avulla. Kiderakenteen perusteella ollaan voitu todentaa aikaisemmin oletettu P4H:n reaktiomekanismi (Eriksson et al. 1989, Longbotham et al. 2015). Ihmisen C-P4H:n kiderakenteen ratkaisun tueksi on julkaistu kiderakenteita muista organismeista kuten Basicillus anthraxisesta C-P4H homodimeeri, jolla on 30%:n homologisuus ihmisen C- P4Hα(I):en nähden (Culpepper et al. 2010). Uusin esitetty olemassa oleva C-P4H rakenne on niin ikään Bascillus anthraxisesta (Schnicker & Dey 2016). Prokollageenien hydroksylaatio on välttämätöntä tropokollageenin biosynteesille. Kaikkien selkärankaisten C-P4H:t ovat α2β2 tetrameereja. Erillinen katalyyttinen domeeni on sitoutunut α- alayksikköön ja substraattia sitova domeeni α- ja β-alayksikön väliin. Kahta αβ heterodimeeriä yhdistää loop-alue (Kivirikko & Myllyharju. 1998). Tetrameerirakenteessa esiintyy viisi konservoitunutta kysteiiniä. Yleensä proteiinissa esiintyvät rikkisillat voivat olla merkittäviä proteiinin aktiivisuudelle ja konformaatiolle. C-P4H:n disulfidisidokset sijoittuvat α-alayksiköön. Disulfidisidokset muodostuvat toisen ja kolmannen, sekä neljännen ja viidennen kysteiinin välille. Tetrameerissa esiintyy kaksi potentiaalista N-glykolysoitua 11
15 motiivia, jotka sijaitsevat α-domeenissa. Toistaiseksi niiden vaikutusta proteiinin liukoisuuteen tai katalyyttiseen aktiivisuuteen ei ole havaittu (Hanauske-Abel & Günzler 1982, John & Bulleid 1996, Lamberg et al. 1995, Levine et al. 1981).Kokopitkää ihmisen C-P4H kristallografiallista rakennetta ei ole vielä ratkaistu. Katalyyttisen α-domeenin, β-domeenin eli PDI:n, peptidi-substraattia sitovan domeenin rakenteita on selvitetty ihmisen C-P4H:sta (Anantharajan et al. 2013, Pekkala et al. 2004) CAT-domeeni osana α-alayksikköä C-P4N:n katalyyttinen domeiini C-terminaalisessa osassa koostuu kolmesta konservoituneesta aminohaposta: Histidiinistä, aspartaatista ja toisesta histidiinistä, joka ulottuu hieman kauemmaksi edellisistä. His-X-Asp-motiivi kelatoi Fe 2+ -kationin. 2-oksoglutaraatti sitoutuu katalyyttisen domeeniin kahdella tavalla. 2-oksoglutaraatti muodostaa ionisidoksen 5-karboksylaattiryhmän Lys 493 :n välillä. Toinen karboksyylipää 2-oksoglutaraatista liittyy yhdeksi Fe 2+ -ionin ligandihampaaksi karbonyylihapen avulla (Majamaa et al. 1984, Majamaa et al. 1986). Neljännen ligandinhampaan Fe 2+ :n kanssa muodostaa molekulaarinen happi. Kompleksin läheisyydessä hieman taaempana on ylimääräinen histidiini, joka ei osallistu katalyysin. Se voi vaikuttaa 2-oksoglutaraatin C 1 -hiilen ja Fe 2+ :n ligandin muodostuksessa ja siten vaikuttaa 2- oksoglutaraatin dekarboksylaatioon. Ihmisen C-P4H:ssa nämä aminohapot ovat His 412, Asp 414 ja His 483. Katalyysiin osallistumaton histidiini on His 501. Myös C-P4H:n katalyyttisessä domeenissa esiintyy kaksoisjuosteinen jelly-roll-laskostyyppi, joka koostuu kahdeksasta anta-paralleelistä β- laskoksesta. Huolimatta useiden eri homologien ratkaisuista kiderakenteista, ihmisen C-P4H:n määrittäminen on usein epäonnistunut kiteytymisongelmien takia (Majamaa et al. 1984, Majamaa et al. 1986, Myllyharju & Kivirikko 1997). C-P4H:n aktiivisuuden mittaus voidaan suorittaa radioaktiivisen 2-oksoglutaraatin avulla, jossa on C 14 -radionuklidi. Reaktiossa voidaan käyttää myös synteettisiä PPG-substraatteja. Aktiivisuus määritetään radionuklidin tuottaman signaalin perusteella. Radionuklidin käyttäminen on menetelmänä vanha (Peterkofsky & Udenfriend 1965), (Hutton et al. 1966)Sen tueksi on kehitelty useita uudempia menetelmiä, joissa radionuklidin käyttöä ei tarvita (Gorres & Raines 2009). 12
16 5.1.2 PSB-domeeni osana N-terminaalista domeenia C-terminaalisen domeenin ohella toinen α-alayksikköön kuuluva N-terminaalinen domeeni käsittää sen alueen α-alayksiköstä, johon substraatti sitoutuu. Tämä osa ihmisen C-P4Hα(I):n kiderakenteesta (aminohapot 1-244, joista substraattia sitova osa ) on ratkaistu (Anantharajan et al. 2013). Siihen kuuluu α2-alayksikköön liittyvä neljästä α-kierteestä rakentuva coiled-coil-laskos, joka on yleisesti oligomeroituvissa proteiineissa esiintyvä laskostyyppi. Substraattina toimivan prokollageeni sitoutuu N-terminaalissa olevaan erilliseen substraattia sitovaan domeeniin (Peptide Substrate Binding domain). PSB:n ja neljän α-kierteen muodostaman coiled-coil laskosta yhdistää löyhä silmukka. PSB:ssä on kaksi TPR-motiivia (Tetratricopeptide repeating domain), jotka toimivat erityisesti proteiini-proteiini/peptidi interaktioissa. C-P4H:n PSB-domeenissa TRP-motiiveja on kaksi, TRP-1 ja TRP-22. Viides α-kierre PSB:ssä on solvatoiva kierre. Toista ja kolmatta kierrettä yhdistää silmukka. Yhdessä TRP-domeenissa on kaksi anti-paralleelia α-kierrettä, joissa kussakin on lähekkäin pieniä, sekä isokokoisia ja hydrofobisia aminohappoja. Esimerkiksi C-P4Hα(I):n TRP-1:n on muotoa Cys 150 -Leu 153 -Gly 154 -Ala 157 -Thr 166 -Met 170 -Ala 173 -Asp 178 (Pekkala et al. 2004). Substraatin liittymisen kannalta tärkeitä tekijöitä PBS:n rakenteessa ovat proteiinin pinnalla olevat tyrosiinitähteet, joita on kahdeksan. Ne sijoittuvat PSB:n koveralle puolelle paitsi Tyr 199 ja Tyr 158, jotka osoittavat poispäin aromaattisesta urasta ja samalla oletetusta peptidisubstraatin sitoutumiskohdasta. Aromaattisen uran tähteistä Tyr 193, Tyr 196 ja Tyr 233 ovat kriittisiä peptidisubstraatin sitoutumisen kannalta. Nämä aminohapot sijoittuvat PSB:n kolmanteen α- kierteeseen. PSB:n aromaattisen uran orientaatio on hiukan erilainen C-P4H:n eri isomuotojen (I,II,III) välillä, joka selittää niiden erilaisen sitoutumisen potentteihin inhibiittoreihin. Lisäksi synteettisten Pro-Pro-Gly-peptidisubstraattien sitoutumisessa on eroja eri isomuotojen välillä (Pekkala et al. 2004) β-alayksikkö-proteiini disulfidi isomeraasi Proteiini disulfidi isomeraasi on tioredoksioksidaasi, joka on saperoniproteiini. Se lokalisoituu ER:n luumeniin, jossa se katalysoi eri proteiinien disulfidisidosten muodostusta, niiden pelkistymistä tai isomerisaatiota (Freedman 1989). Ihmisellä PDI:tä ja C-P4H:ta koodaavat sama geeni. β-alayksikön tunnistaminen PDI:ksi tapahtui havaitsemalla huomattavan suuri identtisyys β-alayksikön ja rotan PDI:tä koodaavalla geenillä (Edman et al. 1985, Pihlajaniemi et al. 1987). 13
17 Ihmisen PDI koostuu 491:stä aminohaposta, jossa on 17:a aminohapon signaalisekvenssi. Sillä on kaksi aktiivista motiivia, jotka sijoittuvat a- ja a -domeeneihin. a- ja b-domeenit sisältävät tioredoksiini laskoksen. Lisäksi b - ja a -domeenien välillä on lyhyt silmukka, mutta sillä ei ole katalyysin kannalta olennaista roolia. C-P4H:n α-alayksikkö liittyy oletettavasti PDI:n a- b - ja a domeeneihin kolmesta eri kohtaa toisistaan etäällä olevien aminohappojen kanssa, jotka ovat His 412, His 483 ja His 501 (Lamberg et al. 1995). Muita tyypillisiä sitoutumisen kannalta tärkeitä tähteitä PDI:ssä on poolittomat leusiinit ja isoleusiinit (Koivunen et al. 2005).Kummassakin katalyyttisessä yksikössä on yksi Cys-Gly-His-Cys-motiivi, jotka toimivat PDI:n isomeraasiaktiivisuuden katalyyttisenä motiivina (Tasanen et al. 1988). C-P4H:den β-domeenin rakenne on identtinen saperoniproteiini PDI:n kanssa (Koivu et al. 1987). PDI:n sitoutuminen α-domeeniin estää sitä agrekoitumasta, pitämällä α-domeenin liukoisessa muodossa. Siten PDI:n sitoutuminen on välttämätöntä α-domeenin katalyyttisen aktiivisuuden kannalta (Vuori et al. 1992). 5.2 Proliinin hydroksylaation vaikutukset tropokollageeniin Niitä proliineja, jotka eivät esiinny X-Y-Gly jaksossa, ei hydroksyloida. Proliinin konformaatio tropokollageeniketjussa on usein (2S) ja hydroksiproliinin (2S,4R). Polaarisen hydroksyyliryhmän elektronegatiivisuudesta johtuen, proliinin hydoroksyloinnilla on vähentävä vaikutus peptidisidoksen resonanssiin (Gorres et al. 2008). Hydrokslyyliryhmän additio pyrrolidiinirenkaaseen muuttaa molekyylin konformaatiota. Vierekkäisten poolisten C γ -C δ1 ja C δ2 -N atomien välinen sidoskulma on 60 astetta, joka on suurin mahdollinen sallittu kulma. Konformaatiota, jossa substituenttien väliset sidokset ovat 60:n asteen kulmassa, kutsutaan ns. gauche efektiksi. Sidoskulmat ohjaavat hydroksiproliinin rengaskonformaation C γ -endo asemaan ja proliinin rengaskonformaation C γ -ekso asemaan. Kollageenien välinen peptidisidos suosii ainoastaan trans konformeeriä (Gorres et al. 2008, Vitagliano et al. 2001). Suotuisin prokollageenin konfromaatio suhteessa C-P4H:hon katalyysivaiheessa on β-käännöslaskos, jolloin sen rakenne on hieman avonaisempi. Katalyysin jälkeen hydroksyyliryhmän vuoksi ketju jäykistyy, eikä sillä ole kovin paljon vapaata tilaa aminohappojen välillä (Chopra & Ananthanarayanan 1982). 14
18 6. Transmembraaninen prolyyli-4-hydroksylaasi 6.1 P4H-TM:n rakenne ja lokalisoituminen C-P4H:n ja HIF-P4H:n lisäksi solulimakalvostossa (ER) esiintyy uudenlainen P4H-entsyymi. Esiintyvä on P4H-TM, joka kuuluu myös Fe(II)-2-oksoglutaraatti dioksigenaasien superperheeseen (Oehme et al. 2002). Poiketen edellisistä P4H-entsyymeistä, P4H-TM sisältää transmembraanisen domeenin, joka sijoittuu oletettavasti aminohappojen välille. P4H-TM:ää ekspressoituu etenkin munuaisissa. Muilta selkärankaisilta P4H-TM geeni löytyy ainakin seeprakalalta (Hyvärinen et al. 2010). P4H-TM:n sekvenssi on konservoitunut hieman suhteessa HIF-P4H:n ja C-P4H:n geeneihin. Sillä on enemmän identtisyyttä C-P4H:den kuin HIF-P4H:den kanssa. Sen aktiivisuus ja reaktion kannalta tärkeät aminohapot vastaavat C-P4H:ta, mutta kuitenkin sen toiminnallisuus liittyy HIF1α:n säätelyyn. P4H-TM ei hydroksyloi kollageenissa esiintyviä proliineja. Sekvenssianalyysien perusteella P4H-TM geeni on 14-15%:sti identtinen ihmisen C-P4Hα(I)- ja α(iii) domeenien kanssa. HIF-P4H 1-3 isomuotojen kanssa P4H-TM:llä on 12-13%:n identtisyys. P4H-TM ei kuitenkaan sisällä samanlaista substraattia sitovaa domeenia (PSB) (Koivunen et al. 2007). P4H-TM:n ekspressio indusoituu HIF1α:n ja HIF2α:n tavoin hypoksiassa ja sen on huomattu rajoittavan HIF2α:n määrää. P4H-TM voi hydroksyloida proliineja HIF1α:n ODDD:stä. HIF1α:n ekspressio indusoituu huomattavasti eniten HIF-P4H 2:n ollessa hiljennettynä, mutta lähes samankaltainen vaikutus on P4H-TM:llä. Siten sillä on samanlainen vaikutus HIF1α:n stabiliteettiin kuin muilla HIF-P4H:lla ja siten sillä on rooli HIF-ohjautuvien geenien säätelyssä (Koivunen et al. 2007). Stoikiometrialtaan P4H-TM on homodimeeri, jolla on kaksi potentiaalista N-glykolysoitua motiivia Asn 368 -Val 369 -Thr 370 ja Asn 382 -Arg 383 -Thr 384. Sen N-terminaalinen osa eroaa muista P4H:sta, mutta sen rautaa ja 2-oksoglutaraattia sitovat aminohapot ovat konservoituneita. P4H-TM:ssä 2- oksoglutaraattia sitoo lysiini. Lysiini esiintyy myös C-P4H:ssa, mutta HIF-P4H:ssa sama aminohappo on arginiini. P4H-TM sisältää EF-hand-domeenin joka sitoo Ca 2+ -ionin Se sijaitsee N-terminaalisessa osassa aminohappojen 192 ja 249 välissä. P4H-TM:n katalyyttinen domeeni suuntautuu luumenin puolelle (Koivunen et al. 2007). 15
19 6.2 P4H-TM:llä ei ole vielä spesifistä roolia P4H-TM:n aktiivisuus on osoittautunut kokeellisissa aktiivisuusmittauksissa heikommaksi kuin HIFp4H:t tai C-P4H:t Kokeellisesti näiden entsyymien aktiivisuutta voidaan mitata epäsuorasti 2- oksoglutaraatin dekarboksylaatioreaktiolla, jossa muodostuu sukkinaattia ja hiilidioksidia. Toistaiseksi P4H-TM:n rooli on läheisesti yhteydessä HIF-signalointiin, mutta sen toiminnasta on saatu lukuisia havaintoja eri kudoksissa. (Myllyharju & Koivunen 2013) Epäsuora aktiivisuuden mittaus voidaan suorittaa ilman substraattia. On huomioitava, että P4H- TM:lle ei ole voitu osoittaa substraattia. Reaktiota seurataan C 14 - tai H 3 -radion(rw.error - Unable to find reference:3)uklidilla leimatun 2-oksoglutaraatista vapautuvan hiilidioksidin avulla. Suurin aktiivisuus esiintyy C-P4H:lla n. 3,0mol/mol entsyymiä/min kun taas P4H-TM sama arvo on vain n.0,2. Tämä johtuu osaltaan P4H-TM ongelmista in vitro olosuhteissa, joka on siten vaikeuttanut karakterisointia (Koivunen et al. 2007). Kirjallisuudesta ei löydy viitteitä sille, että proteiinia oltaisiin onnistuttu ekspressoimaan ja puhdistamaan siinä määrin, jotta sille oltaisiin voitu tehdä kristallografisia määrityksiä. Toisaalta HIF-P4H2:n (McDonough et al. 2006) ja C-P4H α2β2-tetrameerin eri domeenien (Pekkala et al. 2004, (Anantharajan et al. 2013) olemassa olevat kiderakenteet helpottavat P4H-TM:n kiderakenteen selvittämistä. Transmembraanisen osan irtoaminen Asp 88 kohdalta osoittaa proteiinin olevan homodimeeri ja toisaalta helposti hajoava. Toistaiseksi HIF-P4H:lle ei ole pystytty osoittamaan varmasti spesifistä substraattia. Kuitenkin HIF1α:n ODDD proliinit voidaan hydroksyloida P4H:n toimesta, mutta tähän liittyy epävarmuuksia johtuen yllä mainituista in vitro tekijöistä. P 402 :n ja P 564 :n mutaatioilla on kuitenkin havaittu, että P4H-TM voi hydroksyloida sellaisia proliineja ODDD:sta, joita HIF-P4H:t eivät hydroksyloi. (Koivunen et al. 2007). Tämän hetkisten uusimpien tutkimusten mukaan P4H- TM:n geenin puuttumisella tai poikkeavuudella on havaittu muutoksia silmän retinassa ja renaalisessa kudoksessa (Leinonen et al. 2016). 16
20 7. Kirjallisuusluettelo Anantharajan J, Koski M, Kursula P, Hieta R, Bergmann U, Myllyharju J & Wierenga R (2013) The structural motifs for substrate binding and dimerization of the α subunit of collagen prolyl 4-hydroxylase. Structure 21(12): Annunen P, Helaakoski T, Myllyharju J, Veijola J, Pihlajaniemi T & Kivirikko KI (1997) Cloning of the human prolyl 4-hydroxylase alpha subunit isoform alpha(ii) and characterization of the type II enzyme tetramer. The alpha(i) and alpha(ii) subunits do not form a mixed alpha(i)alpha(ii)beta2 tetramer. J Biol Chem. 272(28): Beck I, Ramirez S, Weinmann R & Caro J (1991) Enhancer element at the 3 -flanking region controls transcriptional response to hypoxia in the human erythropoietin gene. J Biol Chem 266(24): Bishop T & Ratcliffe PJ (2014) Signaling hypoxia by hypoxia-inducible factor protein hydroxylases: a historical overview and future perspectives. Hypoxia (Auckl). 2: Bowes JH & Kenten RH (1948) The amino-acid composition and titration curve of collagen. Biochem J. 43(3): Chopra RK & Ananthanarayanan VS (1982) Conformational implications of enzymatic proline hydroxylation in collagen. Proc Natl Acad Sci U S A. 79(23): Compernolle V, Brusselmans K, Acker T, Hoet P, Tjwa M, Beck H, Plaisance S, Dor Y, Keshet E, Lupu F, Nemery B, Dewerchin M, Van Veldhoven P, Plate K, Moons L, Collen D & Carmeliet P (2002) Loss of HIF-2alpha and inhibition of VEGF impair fetal lung maturation, whereas treatment with VEGF prevents fatal respiratory distress in premature mice. Nat Med. 8(7): Costas M, Mehn MP, Jensen MP & Que LJ (2004) Dioxygen activation at mononuclear nonheme iron active sites: enzymes, models, and intermediate. Chem Rev. 104(2): Culpepper MA, Scott EE & Limburg J (2010) Crystal structure of prolyl 4-hydroxylase from Bacillus anthracis. Biochemistry 49(1): de Jong L, Albracht SP & Kemp A (1982) Prolyl 4-hydroxylase activity in relation to the oxidation state of enzyme-bound iron. The role of ascorbate in peptidyl proline hydroxylation. Biochim Biophys Acta. 704(2): Edman JC, Ellis L, Blacher RW, Roth RA & Rutter WJ (1985) Sequence of protein disulphide isomerase and implications of its relationship to thioredoxin. Nature 317(6034): Epstein AC, Gleadle JM, McNeill LA, Hewitson KS, O'Rourke J, Mole DR, Mukherji M, Metzen E, Wilson MI, Dhanda A, Tian YM, Masson N, Hamilton DL, Jaakkola P, Barstead R, Hodgkin J, Maxwell PH, Pugh CW, Schofield CJ & Ratcliffe PJ (2001) C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell. 107(1):
21 Eriksson M, Myllyharju J, Tu H, Hellman M & Kivirikko KI (1989) Evidence for 4-hydroxyproline in viral proteins. Characterization of a viral prolyl 4-hydroxylase and its peptide substrates. Proc Natl Acad Sci U S A. 86(12)): Fischer E (1902) Ber. them. Ges. 36: Freedman SJ, Sun ZY, Poy F, Kung AL, Livingston DM, Wagner G & Eck MJ (2002) Structural basis for recruitment of CBP/p300 by hypoxia-inducible factor-1 alpha. Proc Natl Acad Sci U S A. 99(8): Gorres KL, Edupuganti R, Krow GR & Raines RT (2008) Conformational preferences of substrates for human prolyl 4-hydroxylase. Biochemistry 47(36): Gorres KL & Raines RT (2010) Prolyl-4-hydroxylase. Crit Rev Biochem Mol Biol. 45(2): Gu YZ, Moran SM, Hogenech JB, Wartman L & Bradfield CA (1998) Molecular characterization and chromosomal localization of a third alpha-class hypoxia inducible factor subunit, HIF3alpha. Gene Expr. 7: Hanauske-Abel HM & Günzler V (1982) A stereochemical concept for the catalytic mechanism of prolylhydroxylase: applicability to classification and design of inhibitors. J Theor Biol. 94(2): Heino J & Vuento M (2009) Kollageenit muodostavat soluväliaineen suuret säikeet. In: Anu Stormi (ed) Biokemian ja solubiologian perusteet. Helsinki, WSOY: Helaakoski T, Vuori K, Myllylä R, Kivirikko KI & Pihlajaniemi T (1989) Molecular cloning of the alpha-subunit of human prolyl 4-hydroxylase: the complete cdna-derived amino acid sequence and evidence for alternative splicing of RNA transcripts. Proc Natl Acad Sci U S A. 86(12): Hieta R & Myllyharju J (2002) Cloning and characterization of a low molecular weight prolyl 4- hydroxylase from Arabidopsis thaliana. Effective hydroxylation of proline-rich, collagen-like, and hypoxia-inducible transcription factor alpha-like peptides. J Biol Chem. 277(26): Hoffart LM, Barr EW, Guyer RB, Bollinger JMJ & Krebs C (2006) Direct spectroscopic detection of a C-H-cleaving high-spin Fe(IV) complex in a prolyl-4- hydroxylase. Proc Natl Acad Sci U S A. 103(40): Hoffman EC, Reyes H, Chu FF, Sander H, Conley LH, Brooks BA & Hankinson O (1991) Cloning of a factorrequired for activity of the Ah (dioxin) receptor. Science 252: Huang J, Zhao Q, Mooney SM & Lee FS (2002) Sequence determinants in hypoxia-inducible factor-1alpha for hydroxylation by the prolyl hydroxylases PHD1, PHD2, and PHD3. J Biol Chem. 277(42):
22 Huang LE, Gu J, Schau M & Bunn HF (1998) Regulation of hypoxia-inducible factor 1alpha is mediated by an O2-dependent degradation domain via the ubiquitin-proteasome pathway. Proc Natl Acad Sci U S A. 95(14): Hutton JJ,Jr, Trappel AL & Udenfriend S (1966) A rapid assay for collagen proline hydroxylase. Anal. Biochem. 16: Hyvärinen J, Parikka M, Sormunen R, Rämet M, Tryggvason K, Kivirikko KI, Myllyharju J & Koivunen P (2010) Deficiency of a transmembrane prolyl 4-hydroxylase in the zebrafish leads to basement membrane defects and compromised kidney function. J Biol Chem. 285(53): Ivan M, Haberberger T, Gervasi DC, Michelson KS, Günzler V, Kondo K, Yang H, Sorokina I, Conaway RC, Conaway JW & Kaelin WG (2002) Biochemical purification and pharmacological inhibition of a mammalian prolyl hydroxylase acting on hypoxia-inducible factor. Proc Natl Acad Sci U S A. 99(21): Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, Salic A, Asara JM, Lane WS & Kaelin WG (2001) HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science 292(5516): Iyer NV, Kotch LE, Agani F, Leung SW, Laughner E, Wenger RH, Gassmann M, Gearhart JD, Lawler AM, Yu AY & Semenza GL (1998) Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1 alpha. Genes Dev. 12(2): Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, von Kriegsheim A, Hebestreit HF, Mukherji M, Schofield CJ, Maxwell PH, Pugh CW & Ratcliffe PJ (2001) Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 292(5516): Jiang BH, Rue E, Wang GL, Roe R & Semenza GL (1996) Dimerization, DNA binding, and transactivation properties of hypoxia-inducible factor 1. J Biol Chem 271: John DC & Bulleid NJ (1996) Intracellular dissociation and reassembly of prolyl 4-hydroxylase:the alpha-subunits associated with the immunoglobulin-heavy-chain binding protein (BiP) allowing reassembly with the beta-subunit. Biochem J. 317(3): Kallio PJ, Okamoto K, O'Brien S, Carrero P, Makino Y, Tanaka H & Poellinger L (1998) Signal transduction in hypoxic cells: inducible nuclear translocation and recruitment of the CBP/p300 coactivator by the hypoxia-inducible factor-1alpha. EMBO J. 17(22): Kibel A, Iliopoulos O, DeCaprio JA & Kaelin WGJ (1995) Binding of the von Hippel-Lindau tumor suppressor protein to Elongin B and C. Science 269(5229): Kivirikko KI & Pihlaniemi T (1998) Collagen hydroxylases and the protein disulfide isomerase subunit of prolyl 4-hydroxylases. Adv Enzymol Relat Areas Mol Biol. 72: Kivirikko KI & Prockop DJ (1967) Partial characterization of protocollagen from embryonic cartilage. Biochem J. 102(2):
23 Koivu J, Myllylä R, Helaakoski T, Pihlajaniemi T, Tasanen K & Kivirikko KI (1987) A single polypeptide acts both as the beta subunit of prolyl 4-hydroxylase and as a protein disulfideisomerase. J Biol Chem. 262(14): Koivunen P, Salo KE, Myllyharju J & Ruddock LW (2005) Three binding sites in protein-disulfide isomerase cooperate in collagen prolyl 4-hydroxylase tetramer assembly. J Biol Chem. 280(7): Koivunen P, Tiainen P, Hyvärinen J, Williams KE, Sormunen R, Klaus SJ, Kivirikko KI & Myllyharju J (2007) An endoplasmic reticulum transmembrane prolyl 4-hydroxylase is induced by hypoxia and acts on hypoxia-inducible factor alpha. J Biol Chem. 282(42): Koski MK, Hieta R, Hirsilä M, Rönkä A, Myllyharju J & Wierenga RK (2009) The crystal structure of an algal prolyl 4-hydroxylase complexed with a proline-rich peptide reveals a novel buried tripeptide binding motif. J Biol Chem (37): Krebs C, Galonić-Fujimori D, Walsh CT & Bollinger JMJ (2007) Non-heme Fe(IV)-oxo intermediates. Acc Chem Res. 40(7): Kung AL, Wang S, Klco JM, Kaelin WG & Livingston DM (2000) Suppression of tumor growth through disruption of hypoxia-inducible transcription. Nat. Med 6: Lamberg A, Pihlajaniemi T & Kivirikko KI (1995) Site-directed mutagenesis of the alpha subunit of human prolyl 4-hydroxylase. Identification of three histidine residues critical for catalytic activity. J Biol Chem. 270(17): Lando D, Peet DJ, Whelan DA, Gorman JJ & Whitelaw ML (2002) Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science 295(5556): Leinonen H, Rossi M, Salo AM, Tiainen P, Hyvärinen J, Pitkänen M, Sormunen R, Miinalainen I, Zhang C, Soininen R, Kivirikko KI, Koskelainen A, Tanila H, Myllyharju J & Koivunen P (2016) Lack of P4H-TM in mice results in age-related retinal and renal alterations. Hum Mol Genet. Levine RL (1983) Oxidative modification of glutamine synthetase. I. Inactivation is due to loss of one histidine residue. J Biol Chem. 258(19): Levine RL, Oliver CN, Fulks RM & Stadtman ER (1981) Turnover of bacterial glutamine synthetase: oxidative inactivation precedes proteolysis. Proc Natl Acad Sci U S A. 78(4): Lin F, Suggs S, Lin C, Browne J, Smalling R, Egrie J, Chen K, Fox G, Martin F & Stabinsky Z (1985) Cloning and expression of the human erythropoietin gene. Proc Natl Acad Sci U S A 82(22):
24 Lodish H, Berk A, Kaiser CA, Krieger M, Scott MP, Bretscher A, Ploeg H & Matsudaira P (2007) Integrating cells to tissues. In: Anonymous Molecular biology of the cell. New York, W.H Freeman company: Longbotham JE, Levy C, Johannissen LO, Tarhonskaya H, Jiang S, Loenarz C, Flashman E, Hay S, Schofield S,J & Scrutton NS (2015) Structure and Mechanism of a Viral Collagen Prolyl Hydroxylase. Biochemistry 54(39): Maes C, Carmeliet G & Schipani E (2012) Hypoxia-driven pathways in bone development, regeneration and disease. Nat Rev Rheumatol. 8(6): Majamaa K, Günzler V, Hanauske-Abel HM, Myllylä R, Myllylä R & Kivirikko KI (1986) Partial identity of the 2-oxoglutarate and ascorbate binding sites of prolyl 4-hydroxylase. J Biol Chem. 261(17): Majamaa K, Hanauske-Abel HM, Günzler V & Kivirikko KI (1984) The 2-oxoglutarate binding site of prolyl 4-hydroxylase. Identification of distinct subsites and evidence for 2-oxoglutarate decarboxylation in a ligand reaction at the enzyme-bound ferrous ion. Eur J Biochem. 138(2): Makino Y, Cao R, Svensson K, Bertilsson G, Asman M, Tanaka H, Cao Y, Berkenstam A & Poellinger L (2001) Inhibitory PAS domain protein is a negative regulator of hypoxia-inducible gene expression.. Nature 414(6863): McDonough MA, Li V, Flashman E, Chowdhury R, Mohr C, Liénard BM, Zondlo J, Oldham NJ, Clifton IJ, Lewis J, McNeill LA, Kurzeja RJ, Hewitson KS, Yang E, Jordan S, Syed RS & Schofield CJ (2006) Cellular oxygen sensing: Crystal structure of hypoxia-inducible factor prolyl hydroxylase (PHD2). Proc Natl Acad Sci U S A. 103(26): Myllyharju J & Kivirikko KI (1997) Characterization of the iron- and 2-oxoglutarate-binding sites of human prolyl 4-hydroxylase. EMBO J. 16(6): Myllyharju J & Koivunen P (2013) Hypoxia-inducible factor prolyl 4-hydroxylases: common and specific roles. 394(4): Oehme F, Ellinghaus P, Kolkhof P, Smith TJ, Ramakrishnan S, Hütter J, Schramm M & Flamme I (2002) Overexpression of PH-4, a novel putative proline 4-hydroxylase, modulates activity of hypoxia-inducible transcription factors. Biochem Biophys Res Commun. 296(2): Ohh M, Park CW, Ivan M, Hoffman MA, Kim TY, Huang LE, Pavletich N, Chau V & Kaelin WG (2000) Ubiquitination of hypoxia-inducible factor requires direct binding to the beta-domain of the von Hippel-Lindau protein. Nat Cell Biol. 2(7): Pekkala M, Hieta R, Myllyharju J, Kivirikko K, Wierenga R & Myllyharju J (2004) The peptide-substrate-binding domain of collagen prolyl 4-hydroxylases is a tetratricopeptide repeat domain with functional aromatic residues. J Biol Chem 279(50): Peterkofsky B & Udenfriend S (1965) Enzymatic hydroxylation of proline in microsomal polypeptide leading to formation of collagen. Proc Natl Acad Sci U S A. 53(53):
25 Pihlajaniemi T, Helaakoski T, Tasanen K, Myllylä R, Huhtala ML, Koivu J & Kivirikko KI (1987) Molecular cloning of the beta-subunit of human prolyl 4-hydroxylase. This subunit and protein disulphide isomerase are products of the same gene. EMBO J. 6(3): Rankin EB & Giaccia AJ (2008) The role of hypoxia-inducible factors in tumorigenesis. Cell Death Differ. 15(4): Rankin EB & Giaccia AJ (2016) hypoxic control of metastasis. Science 352(6282): 175. Ryan HE, Lo J & Johnson RS (1998) HIF-1 alpha is required for solid tumor formation and embryonic vascularization. EMBO J. 17(11): Schnicker NJ & Dey M (2016) Bacillus anthracis Prolyl 4-Hydroxylase Modifies Collagen-like Substrates in Asymmetric Patterns. J Biol Chem. 291(25): Semenza GL (2001) HIF-1 and mechanisms of hypoxia sensing. Curr Opin Cell Biol. 13: 167. Semenza GL (2012) Molecular mechanisms mediating metastasis of hypoxic breast cancer cells. Trends Mol Med. 18(9): Shan B, Schaaf C, Schmidt A, Lucia K, Buchfelder M, Losa M, Kuhlen D, Kreutzer J, Perone MJ, Arzt E, Stalla GK & Renner U (2012) Curcumin suppresses HIF1A synthesis and VEGFA release in pituitary adenomas. J Endocrinol. 214(3): Shaw LM & Olsen BR (1991) FACIT collagens: diverse molecular bridges in extracellular matrices. Trends Biochem Sci. 16(5): Stebbins CE, Kaelin WGJ & Pavletich NP (1999) Structure of the VHL-ElonginC-ElonginB complex: implications for VHL tumor suppressor function. Science 284(5413)): Tarhonskaya H, Rydzik AM, Leung IK, Loik ND, Chan MC, Kawamura A, McCullagh JS, Claridge TD, Flashman E & Schofield CJ (2014) Non-enzymatic chemistry enables 2- hydroxyglutarate-mediated activation of 2-oxoglutarate oxygenases. Nat Commun. 5(5): Tasanen K, Parkkonen T, Chow LT, Kivirikko KI & Pihlajaniemi T (1988) Characterization of the human gene for a polypeptide that acts both as the beta subunit of prolyl 4-hydroxylase and as protein disulfide isomerase. J Biol Chem. 263(31): Tian H, McKnight SL & Russell DW (1997) Endothelial PAS domain protein 1 (EPAS1), a transcription factor selectively expressed in endothelial cells. Genes Dev. 11(1): Vitagliano L, Berisio R, Mazzarella L & Zagari A (2001) Structural bases of collagen stabilization induced by proline hydroxylation. Biopolymers 58(5): Vuori K, Pihlajaniemi T, Myllylä R & Kivirikko KI (1992) Site-directed mutagenesis of human protein disulphide isomerase: effect on the assembly, activity and endoplasmic reticulum 22
26 retention of human prolyl 4-hydroxylase in Spodoptera frugiperda insect cells. EMBO J. 11(11)): Wang GL, Jiang BH, Rue E & Semenza GL (1995) Hypoxia-inducible factor 1 is a basic-helixloop-helix-pas heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci U S A. 92(12): Wenger RH & Gassman M (1997) Oxygen(es) and the hypoxia-induc-ible factor-1. Biol. Chem. 378: Yamashita T, Ohneda O, Nagano M, Iemitsu M, Makino Y, Tanaka H, Miyauchi T, Goto K, Ohneda K, Fujii-Kuriyama Y, Poellinger L & Yamamoto M (2008) Abnormal Heart Development and Lung Remodeling in Mice Lackingthe Hypoxia-Inducible Factor-Related Basic Helix-Loop-Helix PAS Protein NEPAS. Mol Cell Biol. 4:


27
ENTSYYMIKATA- LYYSIN PERUSTEET (dos. Tuomas Haltia)
") ENTSYYMIKATA- LYYSIN PERUSTEET (dos. Tuomas Haltia) Elämän edellytykset: Solun täytyy pystyä (a) replikoitumaan (B) katalysoimaan tarvitsemiaan reaktioita tehokkaasti ja selektiivisesti eli sillä on oltava
ENTSYYMIKATA- LYYSIN PERUSTEET (dos. Tuomas Haltia) Elämän edellytykset: Solun täytyy pystyä (a) replikoitumaan (B) katalysoimaan tarvitsemiaan reaktioita tehokkaasti ja selektiivisesti eli sillä on oltava
Peptidi ---- F ----- K ----- V ----- R ----- H ----- A ---- A. Siirtäjä-RNA:n (trna:n) (3 ) AAG UUC CAC GCA GUG CGU (5 ) antikodonit
 (3 ) AAG UUC CAC GCA GUG CGU (5 ) antikodonit") Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 3 Pisteet / 20 Osa 1: Haluat selvittää -- F -- K -- V -- R -- H -- A peptidiä
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 24.5.2006 Etunimet Tehtävä 3 Pisteet / 20 Osa 1: Haluat selvittää -- F -- K -- V -- R -- H -- A peptidiä
Vastaa lyhyesti selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1 1) Tunnista molekyylit (1 piste) ja täytä seuraava taulukko (2 pistettä) a) b) c) d) a) Syklinen AMP (camp) (0.25) b) Beta-karoteeni (0.25 p) c) Sakkaroosi (0.25 p) d) -D-Glukopyranoosi (0.25 p) 2 Taulukko.
1 1) Tunnista molekyylit (1 piste) ja täytä seuraava taulukko (2 pistettä) a) b) c) d) a) Syklinen AMP (camp) (0.25) b) Beta-karoteeni (0.25 p) c) Sakkaroosi (0.25 p) d) -D-Glukopyranoosi (0.25 p) 2 Taulukko.
Ma > GENERAL PRINCIPLES OF CELL SIGNALING
 Ma 5.12. -> GENERAL PRINCIPLES OF CELL SIGNALING Cell-Surface Receptors Relay Extracellular Signals via Intracellular Signaling Pathways Some Intracellular Signaling Proteins Act as Molecular Switches
Ma 5.12. -> GENERAL PRINCIPLES OF CELL SIGNALING Cell-Surface Receptors Relay Extracellular Signals via Intracellular Signaling Pathways Some Intracellular Signaling Proteins Act as Molecular Switches
Biokemian perusteet 26.9.2012: Hemoglobiini, Entsyymikatalyysi
 Biokemian perusteet 26.9.2012: Hemoglobiini, Entsyymikatalyysi Dos. Tuomas Haltia Sirppisoluanemia, Hb-mutaatio Glu-6 Val Hemoglobiini allosteerinen hapen kuljettajaproteiini (ei ole entsyymi!) Allosteerinen
Biokemian perusteet 26.9.2012: Hemoglobiini, Entsyymikatalyysi Dos. Tuomas Haltia Sirppisoluanemia, Hb-mutaatio Glu-6 Val Hemoglobiini allosteerinen hapen kuljettajaproteiini (ei ole entsyymi!) Allosteerinen
Bioteknologian tutkinto-ohjelma Valintakoe Tehtävä 3 Pisteet / 30
 Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 3 Pisteet / 30 3. a) Alla on lyhyt jakso dsdna:ta, joka koodaa muutaman aminohappotähteen
Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 3 Pisteet / 30 3. a) Alla on lyhyt jakso dsdna:ta, joka koodaa muutaman aminohappotähteen
PROTEIINIEN MUOKKAUS JA KULJETUS
 PROTEIINIEN MUOKKAUS JA KULJETUS 1.1 Endoplasmakalvosto Endoplasmakalvosto on organelli joka sijaitsee tumakalvossa kiinni. Se on topologisesti siis yhtä tumakotelon kanssa. Se koostuu kahdesta osasta:
PROTEIINIEN MUOKKAUS JA KULJETUS 1.1 Endoplasmakalvosto Endoplasmakalvosto on organelli joka sijaitsee tumakalvossa kiinni. Se on topologisesti siis yhtä tumakotelon kanssa. Se koostuu kahdesta osasta:
NON-CODING RNA (ncrna)
") NON-CODING RNA (ncrna) 1. Yleistä NcRNA eli non-coding RNA tarkoittaa kaikkia proteiinia koodaamattomia rnamolekyylejä. Näistä yleisimmin tunnetut ovat ribosomaalinen RNA (rrna) sekä siirtäjä-rna (trna),
NON-CODING RNA (ncrna) 1. Yleistä NcRNA eli non-coding RNA tarkoittaa kaikkia proteiinia koodaamattomia rnamolekyylejä. Näistä yleisimmin tunnetut ovat ribosomaalinen RNA (rrna) sekä siirtäjä-rna (trna),
ELEC-C2210 Molekyyli- ja solubiologia
 ELEC-C2210 Molekyyli- ja solubiologia Entsyymikatalyysi Vuento & Heino ss. 66-75 ECB: Luku 3, s. 90-93 & luku 4, s. 144- Dos. Tuomas Haltia, Biotieteiden laitos, biokemia ja biotekniikka Miten entsyymit
ELEC-C2210 Molekyyli- ja solubiologia Entsyymikatalyysi Vuento & Heino ss. 66-75 ECB: Luku 3, s. 90-93 & luku 4, s. 144- Dos. Tuomas Haltia, Biotieteiden laitos, biokemia ja biotekniikka Miten entsyymit
DNA RNA proteiinit transkriptio prosessointi translaatio regulaatio
 CELL 411-- replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi
CELL 411-- replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA proteiinit transkriptio prosessointi
Genomin ilmentyminen Liisa Kauppi, Genomibiologian tutkimusohjelma
 Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Genomin ilmentyminen 17.1.2013 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Genomin ilmentyminen transkription aloitus RNA:n synteesi ja muokkaus DNA:n ja RNA:n välisiä eroja
Biomolekyylit 2. Nukleotidit, aminohapot ja proteiinit
 Biomolekyylit 2 Nukleotidit, aminohapot ja proteiinit Nukleotidit Ihmisen perimä, eli DNA (deoksiribonukleiinihappo) muodostuu pitkästä nukleotidiketjusta. Lisäksi nukleotidit toimivat mm. proteiinisynteesissä
Biomolekyylit 2 Nukleotidit, aminohapot ja proteiinit Nukleotidit Ihmisen perimä, eli DNA (deoksiribonukleiinihappo) muodostuu pitkästä nukleotidiketjusta. Lisäksi nukleotidit toimivat mm. proteiinisynteesissä
Peptidisynteesi. SPPS:n Periaate
 Tapio Nevalainen Lääkeainesynteesit II 2011 eptidisynteesi i eptidisynteesi Suoritetaan yleensä kiinteän faasin pinnalla; solid phase peptide synthesis (SS) Suuret peptidiainemäärät valmistetaan liuosfaasissa.
Tapio Nevalainen Lääkeainesynteesit II 2011 eptidisynteesi i eptidisynteesi Suoritetaan yleensä kiinteän faasin pinnalla; solid phase peptide synthesis (SS) Suuret peptidiainemäärät valmistetaan liuosfaasissa.
Happi solujen toimintojen säätelijänä Mikko Nikinmaa
 Happi solujen toimintojen säätelijänä Mikko Nikinmaa Elämä maapallolla nykymuotoisena olisi mahdotonta ilman happea. Happi osallistuu mitokondrioiden energiantuotantoon vastaanottamalla elektroninsiirtoketjun
Happi solujen toimintojen säätelijänä Mikko Nikinmaa Elämä maapallolla nykymuotoisena olisi mahdotonta ilman happea. Happi osallistuu mitokondrioiden energiantuotantoon vastaanottamalla elektroninsiirtoketjun
Genomi-ilmentyminen Genom expression (uttryckning) Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia
 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia") Genomi-ilmentyminen Genom expression (uttryckning) DNA RNA 7.12.2017 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia Osaamistavoitteet Lärandemål Luennon jälkeen ymmärrät pääperiaatteet
Genomi-ilmentyminen Genom expression (uttryckning) DNA RNA 7.12.2017 Nina Peitsaro, yliopistonlehtori, Medicum, Biokemia ja Kehitysbiologia Osaamistavoitteet Lärandemål Luennon jälkeen ymmärrät pääperiaatteet
Yhtäläisyydet selkärankaisten aivoissa, osa II. Niko Lankinen
 Yhtäläisyydet selkärankaisten aivoissa, osa II Niko Lankinen Sisältö Neuroneille tyypilliset molekyylit Suoraa jatkoa Niinan esitykseen Alkion aivojen vertailua Neuromeerinen malli Neuromeerisen mallin
Yhtäläisyydet selkärankaisten aivoissa, osa II Niko Lankinen Sisältö Neuroneille tyypilliset molekyylit Suoraa jatkoa Niinan esitykseen Alkion aivojen vertailua Neuromeerinen malli Neuromeerisen mallin
Essential Cell Biology
 Alberts Bray Hopkin Johnson Lewis Raff Roberts Walter Essential Cell Biology FOURTH EDITION Chapter 16 Cell Signaling Copyright Garland Science 2014 1 GENERAL PRINCIPLES OF CELL SIGNALING Signals Can Act
Alberts Bray Hopkin Johnson Lewis Raff Roberts Walter Essential Cell Biology FOURTH EDITION Chapter 16 Cell Signaling Copyright Garland Science 2014 1 GENERAL PRINCIPLES OF CELL SIGNALING Signals Can Act
Biomolekyylit ja biomeerit
 Biomolekyylit ja biomeerit Polymeerit ovat hyvin suurikokoisia, pitkäketjuisia molekyylejä, jotka muodostuvat monomeereista joko polyadditio- tai polykondensaatioreaktiolla. Polymeerit Synteettiset polymeerit
Biomolekyylit ja biomeerit Polymeerit ovat hyvin suurikokoisia, pitkäketjuisia molekyylejä, jotka muodostuvat monomeereista joko polyadditio- tai polykondensaatioreaktiolla. Polymeerit Synteettiset polymeerit
Käsitteitä. Hormones and the Endocrine System Hormonit ja sisäeritejärjestelmä. Sisäeriterauhanen
 Käsitteitä Hormones and the Endocrine System Hormonit ja sisäeritejärjestelmä 1/2 Umpirauhanen vs. sisäeriterauhanen Endokrinologia Parakriininen Autokriininen Neurotransmitteri Reseptori Sisäeriterauhanen
Käsitteitä Hormones and the Endocrine System Hormonit ja sisäeritejärjestelmä 1/2 Umpirauhanen vs. sisäeriterauhanen Endokrinologia Parakriininen Autokriininen Neurotransmitteri Reseptori Sisäeriterauhanen
Valmiustaitoja biokemisteille
 Valmiustaitoja biokemisteille - Power Point -ohjelman käyttö - seminaariesitelmän laatiminen ja esittäminen Tuomo Glumoff 1. Power Point -ohjelman käyttö - teksti - kuvat - tausta - valmiit pohjat * löytyy
Valmiustaitoja biokemisteille - Power Point -ohjelman käyttö - seminaariesitelmän laatiminen ja esittäminen Tuomo Glumoff 1. Power Point -ohjelman käyttö - teksti - kuvat - tausta - valmiit pohjat * löytyy
Epigeneettinen säätely ja genomin leimautuminen. Tiina Immonen BLL Biokemia ja kehitysbiologia
 Epigeneettinen säätely ja genomin leimautuminen Tiina Immonen BLL Biokemia ja kehitysbiologia 21.1.2014 Epigeneettinen säätely Epigenetic: may be used for anything to do with development, but nowadays
Epigeneettinen säätely ja genomin leimautuminen Tiina Immonen BLL Biokemia ja kehitysbiologia 21.1.2014 Epigeneettinen säätely Epigenetic: may be used for anything to do with development, but nowadays
Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe Tehtävä 1 Pisteet / 15
 Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
Tampereen yliopisto Henkilötunnus - Lääketieteen ja biotieteiden tiedekunta Sukunimi Bioteknologia tutkinto-ohjelma Etunimet valintakoe pe 18.5.2018 Tehtävä 1 Pisteet / 15 1. Alla on esitetty urheilijan
Supporting Information for
 Supporting Information for Analysis of Sogatella furcifera proteome that interact with P10 protein of Southern rice black-streaked dwarf virus Win Than*, Faliang Qin*, Wenwen Liu, Xifeng Wang ** State
Supporting Information for Analysis of Sogatella furcifera proteome that interact with P10 protein of Southern rice black-streaked dwarf virus Win Than*, Faliang Qin*, Wenwen Liu, Xifeng Wang ** State
Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20
 Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20 3: Osa 1 Tumallisten solujen genomin toiminnassa sekä geenien
Helsingin yliopisto/tampereen yliopisto Henkilötunnus - Biokemian/bioteknologian valintakoe Sukunimi 26. 05. 2005 Etunimet Tehtävä 3 Pisteet / 20 3: Osa 1 Tumallisten solujen genomin toiminnassa sekä geenien
Ota henkilötodistus mukaasi jättäessäsi vastauspaperin. Kysymyksiin vastataan suomeksi.
 Oulun yliopiston biokemian koulutusohjelman valintakoe 20.5.2016 Nimi: Ota henkilötodistus mukaasi jättäessäsi vastauspaperin. Kysymyksiin vastataan suomeksi. Osa 1 Aineistotehtävä. Vastaa vain varattuun
Oulun yliopiston biokemian koulutusohjelman valintakoe 20.5.2016 Nimi: Ota henkilötodistus mukaasi jättäessäsi vastauspaperin. Kysymyksiin vastataan suomeksi. Osa 1 Aineistotehtävä. Vastaa vain varattuun
BI4 IHMISEN BIOLOGIA
 BI4 IHMISEN BIOLOGIA IHMINEN ON TOIMIVA KOKONAISUUS Ihmisessä on noin 60 000 miljardia solua Solujen perusrakenne on samanlainen, mutta ne ovat erilaistuneet hoitamaan omia tehtäviään Solujen on oltava
BI4 IHMISEN BIOLOGIA IHMINEN ON TOIMIVA KOKONAISUUS Ihmisessä on noin 60 000 miljardia solua Solujen perusrakenne on samanlainen, mutta ne ovat erilaistuneet hoitamaan omia tehtäviään Solujen on oltava
VASTAUS 1: Yhdistä oikein
 KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
KPL3 VASTAUS 1: Yhdistä oikein a) haploidi - V) ihmisen sukusolu b) diploidi - IV) ihmisen somaattinen solu c) polyploidi - VI) 5n d) iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen
LIGNIINI yleisesti käytettyjä termejä
 Luennon 9 oppimistavoitteet Ligniinin biosynteesi, rakenne ja ominaisuudet Puu-19210 Puun rakenne ja kemia Ymmärrät, että ligniini on amorfinen makromolekyyli, joka muodostuu monomeeriyksiköistä Tiedät
Luennon 9 oppimistavoitteet Ligniinin biosynteesi, rakenne ja ominaisuudet Puu-19210 Puun rakenne ja kemia Ymmärrät, että ligniini on amorfinen makromolekyyli, joka muodostuu monomeeriyksiköistä Tiedät
Genomin ilmentyminen
 Kauppi 17/01/2014 Genomin ilmentyminen LH1, Molekyylibiologia 17.1.2014 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Huone C501b, Biomedicum 1 Transkriptiofaktorin mutaatio voi
Kauppi 17/01/2014 Genomin ilmentyminen LH1, Molekyylibiologia 17.1.2014 Liisa Kauppi, Genomibiologian tutkimusohjelma liisa.kauppi@helsinki.fi Huone C501b, Biomedicum 1 Transkriptiofaktorin mutaatio voi
Nimi sosiaaliturvatunnus. Vastaa lyhyesti, selkeällä käsialalla. Vain vastausruudun sisällä olevat tekstit, kuvat jne huomioidaan
 1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
1. a) Mitä tarkoitetaan biopolymeerilla? Mihin kolmeen ryhmään biopolymeerit voidaan jakaa? (1,5 p) Biopolymeerit ovat luonnossa esiintyviä / elävien solujen muodostamia polymeerejä / makromolekyylejä.
Epigeneettinen säätely ja genomin leimautuminen. Tiina Immonen Medicum, Biokemia ja kehitysbiologia
 Epigeneettinen säätely ja genomin leimautuminen Tiina Immonen Medicum, Biokemia ja kehitysbiologia 12.12.2017 Epigenetic inheritance: A heritable alteration in a cell s or organism s phenotype that does
Epigeneettinen säätely ja genomin leimautuminen Tiina Immonen Medicum, Biokemia ja kehitysbiologia 12.12.2017 Epigenetic inheritance: A heritable alteration in a cell s or organism s phenotype that does
KEMIA HYVÄN VASTAUKSEN PIIRTEET
 BILÄÄKETIETEEN enkilötunnus: - KULUTUSJELMA Sukunimi: 20.5.2015 Etunimet: Nimikirjoitus: KEMIA Kuulustelu klo 9.00-13.00 YVÄN VASTAUKSEN PIIRTEET Tehtävämonisteen tehtäviin vastataan erilliselle vastausmonisteelle.
BILÄÄKETIETEEN enkilötunnus: - KULUTUSJELMA Sukunimi: 20.5.2015 Etunimet: Nimikirjoitus: KEMIA Kuulustelu klo 9.00-13.00 YVÄN VASTAUKSEN PIIRTEET Tehtävämonisteen tehtäviin vastataan erilliselle vastausmonisteelle.
Avaruus- eli stereoisomeria
 Avaruus- eli stereoisomeria KEMIAN MIKRO- MAAILMA, KE2 Kolme alalajia: 1) cis-trans-isomeria, 2) optinen isomeria ja 3) konformaatioisomeria, Puhtaiden stereoisomeerien valmistaminen ja erottaminen toisistaan
Avaruus- eli stereoisomeria KEMIAN MIKRO- MAAILMA, KE2 Kolme alalajia: 1) cis-trans-isomeria, 2) optinen isomeria ja 3) konformaatioisomeria, Puhtaiden stereoisomeerien valmistaminen ja erottaminen toisistaan
Biopolymeerit. Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä.
 Biopolymeerit Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä. Tärkeimpiä biopolymeerejä ovat hiilihydraatit, proteiinit ja nukleiinihapot. 1 Hiilihydraatit Hiilihydraatit jaetaan mono
Biopolymeerit Biopolymeerit ovat kasveissa ja eläimissä esiintyviä polymeerejä. Tärkeimpiä biopolymeerejä ovat hiilihydraatit, proteiinit ja nukleiinihapot. 1 Hiilihydraatit Hiilihydraatit jaetaan mono
Soluväliaineen tutkimuksen monet ulottuvuudet
 ÄYRÄPÄÄN LUENTO Taina Pihlajaniemi Soluväliaineen tutkimuksen monet ulottuvuudet Soluväliaineella on tärkeä kudoksia tukeva rakenteellinen tehtävänsä, jonka lisäksi se säätelee solujen vasteita ympäristölleen
ÄYRÄPÄÄN LUENTO Taina Pihlajaniemi Soluväliaineen tutkimuksen monet ulottuvuudet Soluväliaineella on tärkeä kudoksia tukeva rakenteellinen tehtävänsä, jonka lisäksi se säätelee solujen vasteita ympäristölleen
Proteiinin rakenteen selvittämisestä ja visualisoinnista
 TKK Solubiosysteemien perusteet syksy 2002 Harkkatyö M.Tarvainen Proteiinin rakenteen selvittämisestä ja visualisoinnista 1. Yleistä proteiineista 2. Röntgensädekristallografia 3. Ydinmagneettinen resonanssimenetelmä
TKK Solubiosysteemien perusteet syksy 2002 Harkkatyö M.Tarvainen Proteiinin rakenteen selvittämisestä ja visualisoinnista 1. Yleistä proteiineista 2. Röntgensädekristallografia 3. Ydinmagneettinen resonanssimenetelmä
DNA:n informaation kulku, koostumus
 DNA:n informaation kulku, koostumus KOOSTUMUS Elävien bio-organismien koostumus. Vety, hiili, happi ja typpi muodostavat yli 99% orgaanisten molekyylien rakenneosista. Biomolekyylit voidaan pääosin jakaa
DNA:n informaation kulku, koostumus KOOSTUMUS Elävien bio-organismien koostumus. Vety, hiili, happi ja typpi muodostavat yli 99% orgaanisten molekyylien rakenneosista. Biomolekyylit voidaan pääosin jakaa
vauriotyypit Figure 5-17.mhc.restriktio 9/24/14 Autoimmuniteetti Kudosvaurion mekanismit Petteri Arstila Haartman-instituutti Patogeeniset mekanismit
 vauriotyypit Kudosvaurion mekanismit Autoimmuniteetti Petteri Arstila Haartman-instituutti Antigeenin tunnistus HLA:ssa pitää sisällään autoimmuniteetin riskin: jokaisella on autoreaktiivisia lymfosyyttejä
vauriotyypit Kudosvaurion mekanismit Autoimmuniteetti Petteri Arstila Haartman-instituutti Antigeenin tunnistus HLA:ssa pitää sisällään autoimmuniteetin riskin: jokaisella on autoreaktiivisia lymfosyyttejä
GLYKOLYYSI! Riikka Kivelä, LitT Tutkijatohtori Wihurin tutkimuslaitos ja Translationaalisen syöpäbiologian tutkimusohjelma Helsingin yliopisto
 GLYKLYYSI! Riikka Kivelä, LitT Tutkijatohtori Wihurin tutkimuslaitos ja Translationaalisen syöpäbiologian tutkimusohjelma Helsingin yliopisto LUENNN RAKENNE! Anaerobinen (hapeton) vs. Aerobinen (hapellinen)
GLYKLYYSI! Riikka Kivelä, LitT Tutkijatohtori Wihurin tutkimuslaitos ja Translationaalisen syöpäbiologian tutkimusohjelma Helsingin yliopisto LUENNN RAKENNE! Anaerobinen (hapeton) vs. Aerobinen (hapellinen)
Luennon 5 oppimistavoitteet. Soluseinän biosynteesi. Puu-19.210 Puun rakenne ja kemia. Solun organelleja. Elävä kasvisolu
 Luennon 5 oppimistavoitteet Soluseinän biosynteesi Puu-19.210 Puun rakenne ja kemia saat listata puuaineksen muodostumisen vaiheet. Ymmärrät, kuinka soluseinän tapahtuu. saat lyhyesti kuvata soluseinän
Luennon 5 oppimistavoitteet Soluseinän biosynteesi Puu-19.210 Puun rakenne ja kemia saat listata puuaineksen muodostumisen vaiheet. Ymmärrät, kuinka soluseinän tapahtuu. saat lyhyesti kuvata soluseinän
HEIKOT VUOROVAIKUTUKSET MOLEKYYLIEN VÄLISET SIDOKSET
 HEIKOT VUOROVAIKUTUKSET MOLEKYYLIEN VÄLISET SIDOKSET Tunnin sisältö 2. Heikot vuorovaikutukset Millaisia erilaisia? Missä esiintyvät? Biologinen/lääketieteellinen merkitys Heikot sidokset Dipoli-dipolisidos
HEIKOT VUOROVAIKUTUKSET MOLEKYYLIEN VÄLISET SIDOKSET Tunnin sisältö 2. Heikot vuorovaikutukset Millaisia erilaisia? Missä esiintyvät? Biologinen/lääketieteellinen merkitys Heikot sidokset Dipoli-dipolisidos
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi
 6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
6 GEENIT OHJAAVAT SOLUN TOIMINTAA nukleiinihapot DNA ja RNA Geenin rakenne Geneettinen informaatio Proteiinisynteesi GENEETTINEN INFORMAATIO Geeneihin pakattu informaatio ohjaa solun toimintaa ja siirtyy
Kovalenttinen sidos ja molekyyliyhdisteiden ominaisuuksia
 Kovalenttinen sidos ja molekyyliyhdisteiden ominaisuuksia 16. helmikuuta 2014/S.. Mikä on kovalenttinen sidos? Kun atomit jakavat ulkoelektronejaan, syntyy kovalenttinen sidos. Kovalenttinen sidos on siis
Kovalenttinen sidos ja molekyyliyhdisteiden ominaisuuksia 16. helmikuuta 2014/S.. Mikä on kovalenttinen sidos? Kun atomit jakavat ulkoelektronejaan, syntyy kovalenttinen sidos. Kovalenttinen sidos on siis
Reseptoripotentiaalista (RP) aktiopotentiaaliin
 aktiopotentiaaliin") Haju- ja makuaisti Reseptoripotentiaalista (RP) aktiopotentiaaliin Reseptoristimulaatio lokaalinen sähköinen ärtyminen (melkein aina depolarisaatio) RP syntymekanismi vaihtelee aistimesta toiseen RP leviää
Haju- ja makuaisti Reseptoripotentiaalista (RP) aktiopotentiaaliin Reseptoristimulaatio lokaalinen sähköinen ärtyminen (melkein aina depolarisaatio) RP syntymekanismi vaihtelee aistimesta toiseen RP leviää
Drosophila on kehitysgenetiikan mallilaji nro 1
 Drosophila on kehitysgenetiikan mallilaji nro 1 replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA
Drosophila on kehitysgenetiikan mallilaji nro 1 replikaatio repair mitoosi meioosi fertilisaatio rekombinaatio repair mendelistinen genetiikka DNA-huusholli Geenien toiminta molekyyligenetiikka DNA RNA
Ribosomit 1. Ribosomit 2. Ribosomit 3
 Ribosomit 1 Palade & Siekevitz eristivät jaottelusentrifugaatiolla ns. mikrosomeja radioakt. aminohapot kertyivät mikrosomeihin, jotka peräisin rer:ää sisältävistä soluista proteiinisynteesi soluliman
Ribosomit 1 Palade & Siekevitz eristivät jaottelusentrifugaatiolla ns. mikrosomeja radioakt. aminohapot kertyivät mikrosomeihin, jotka peräisin rer:ää sisältävistä soluista proteiinisynteesi soluliman
Supporting information
 Supporting information Figure S1. Carotenoid biosynthesis pathway in papaya fruit which is adopted from Blas et al. (2010) (reference 4) and Nisar et al. (2015) (reference 5). Carotenoids are synthesized
Supporting information Figure S1. Carotenoid biosynthesis pathway in papaya fruit which is adopted from Blas et al. (2010) (reference 4) and Nisar et al. (2015) (reference 5). Carotenoids are synthesized
Kuva 1. Korento lehdellä (Miettinen, A., 2006).
.") Pigmentit yönteisten värien monimuotoisuus (ks. kuva 1) on aina kiehtonut luonnontieteilijöitä. Vasta 1900-luvun alussa alettiin ymmärtää pigmenttien kemiaa. Pigmentit esiintyvät luonnon materiaaleissa
Pigmentit yönteisten värien monimuotoisuus (ks. kuva 1) on aina kiehtonut luonnontieteilijöitä. Vasta 1900-luvun alussa alettiin ymmärtää pigmenttien kemiaa. Pigmentit esiintyvät luonnon materiaaleissa
Solun kemiallinen peruskoostumus eläinsolu. Solun kemia. Solun kemiallinen peruskoostumus bakteerisolu. Vesi 1
 Solun kemiallinen peruskoostumus eläinsolu Solun kemia paino-% Vesi 75-90 proteiinit 10-20 Lipidit 2 Hiilihydraatit 1 RNA/DNA 0,7/0,4 Epäorg. 1,5 Solun kemiallinen peruskoostumus bakteerisolu Vesi 1 paino-%
Solun kemiallinen peruskoostumus eläinsolu Solun kemia paino-% Vesi 75-90 proteiinit 10-20 Lipidit 2 Hiilihydraatit 1 RNA/DNA 0,7/0,4 Epäorg. 1,5 Solun kemiallinen peruskoostumus bakteerisolu Vesi 1 paino-%
CHEM-A1200 Kemiallinen rakenne ja sitoutuminen
 CHEM-A1200 Kemiallinen rakenne ja sitoutuminen Hapot, Emäkset ja pk a Opettava tutkija Pekka M Joensuu Jokaisella hapolla on: Arvo, joka kertoo meille kuinka hapan kyseinen protoni on. Helpottaa valitsemaan
CHEM-A1200 Kemiallinen rakenne ja sitoutuminen Hapot, Emäkset ja pk a Opettava tutkija Pekka M Joensuu Jokaisella hapolla on: Arvo, joka kertoo meille kuinka hapan kyseinen protoni on. Helpottaa valitsemaan
Avaruus- eli stereoisomeria
 Avaruus- eli stereoisomeria IHMISEN JA ELINYMPÄ- RISTÖN KEMIAA, KE2 Kolme alalajia: 1) cis-trans-isomeria, 2) optinen isomeria ja 3) konformaatioisomeria, Puhtaiden stereoisomeerien valmistaminen ja erottaminen
Avaruus- eli stereoisomeria IHMISEN JA ELINYMPÄ- RISTÖN KEMIAA, KE2 Kolme alalajia: 1) cis-trans-isomeria, 2) optinen isomeria ja 3) konformaatioisomeria, Puhtaiden stereoisomeerien valmistaminen ja erottaminen
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita
 Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 10. Valkuaisaineiden valmistaminen solussa 1. Avainsanat 2. Perinnöllinen tieto on dna:n emäsjärjestyksessä 3. Proteiinit koostuvat
Perinnöllisyystieteen perusteita III Perinnöllisyystieteen perusteita 10. Valkuaisaineiden valmistaminen solussa 1. Avainsanat 2. Perinnöllinen tieto on dna:n emäsjärjestyksessä 3. Proteiinit koostuvat
Molekyyli- ja solubiologia ELEC-2210 Proteiinit
 Molekyyli- ja solubiologia ELEC-2210 Proteiinit Vuento & Heino: Biokemian ja solubiologian perusteet, ss. 51-66 Alberts et al. Essential Cell Biology, 4. p, luku 4 Dos. Tuomas Haltia, HY, Biotieteiden
Molekyyli- ja solubiologia ELEC-2210 Proteiinit Vuento & Heino: Biokemian ja solubiologian perusteet, ss. 51-66 Alberts et al. Essential Cell Biology, 4. p, luku 4 Dos. Tuomas Haltia, HY, Biotieteiden
Bioteknologian tutkinto-ohjelma. 1. a) Rastita, mitkä seuraavista väittämistä ovat oikein ja mitkä väärin. (10 p.)
 Rastita, mitkä seuraavista väittämistä ovat oikein ja mitkä väärin. (10 p.)") Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 1 Pisteet / 30 1. a) Rastita, mitkä seuraavista väittämistä ovat oikein ja mitkä väärin.
Tampereen yliopisto Bioteknologian tutkinto-ohjelma Valintakoe 21.5.2015 Henkilötunnus - Sukunimi Etunimet Tehtävä 1 Pisteet / 30 1. a) Rastita, mitkä seuraavista väittämistä ovat oikein ja mitkä väärin.
Luun aineenvaihdunnan biokemialliset mittarit: mitä, miksi ja milloin
 Luun aineenvaihdunnan biokemialliset mittarit: mitä, miksi ja milloin Marja-Kaisa Koivula FT, dosentti, sairaalakemisti 22.5.2014 Esityksen sisältö Mitä luun aineenvaihdunnalla tarkoitetaan? Mitä ovat
Luun aineenvaihdunnan biokemialliset mittarit: mitä, miksi ja milloin Marja-Kaisa Koivula FT, dosentti, sairaalakemisti 22.5.2014 Esityksen sisältö Mitä luun aineenvaihdunnalla tarkoitetaan? Mitä ovat
Viral DNA as a model for coil to globule transition
 Viral DNA as a model for coil to globule transition Marina Rossi Lab. of complex fluids and molecular biophysics LITA (Segrate) UNIVERSITA DEGLI STUDI DI MILANO - PhD Workshop October 14 th, 2013 Temperature
Viral DNA as a model for coil to globule transition Marina Rossi Lab. of complex fluids and molecular biophysics LITA (Segrate) UNIVERSITA DEGLI STUDI DI MILANO - PhD Workshop October 14 th, 2013 Temperature
Kurssiin sisältyvät kappaleet kirjasta Tortora et al. (Microbiology) ja Alberts et al. (Essential Cell Biology), 1/2
 ja Alberts et al. (Essential Cell Biology), 1/2") Kurssiin sisältyvät kappaleet kirjasta Tortora et al. (Microbiology) ja Alberts et al. (Essential Cell Biology), 1/2 Luento 1: Yleisten asioiden jälkeen: MB, luku 1 (s. 28-49) Luento 2: Luku 1 jatkuu +
Kurssiin sisältyvät kappaleet kirjasta Tortora et al. (Microbiology) ja Alberts et al. (Essential Cell Biology), 1/2 Luento 1: Yleisten asioiden jälkeen: MB, luku 1 (s. 28-49) Luento 2: Luku 1 jatkuu +
GLYKOLYYSI! Glykolyyttinen metaboliareitti! LUENNON RAKENNE! ENERGIA HIILIHYDRAATEISTA. ATP:n ANAEROBINEN JA AEROBINEN UUDELLEENMUODOSTUS
 GLYKLYYSI LUENNN RAKENNE Anaerobinen (hapeton) vs. Aerobinen (hapellinen) energiantuotto Glykolyysin reaktiot Glykolyysin energetiikka Glykolyysi / anaerobinen energiantuotto syövässä ja liikunnassa Riikka
GLYKLYYSI LUENNN RAKENNE Anaerobinen (hapeton) vs. Aerobinen (hapellinen) energiantuotto Glykolyysin reaktiot Glykolyysin energetiikka Glykolyysi / anaerobinen energiantuotto syövässä ja liikunnassa Riikka
Kipu. Oleg Kambur. Geneettisillä tekijöillä suuri merkitys Yksittäisiä geenejä on löydetty vain vähän COMT 23.6.2015
 Katekoli-O-metyylitransferaasi ja kipu Oleg Kambur Kipu Geneettisillä tekijöillä suuri merkitys Yksittäisiä geenejä on löydetty vain vähän COMT 1 Katekoli-O-metyylitransferaasi (COMT) proteiini tuotetaan
Katekoli-O-metyylitransferaasi ja kipu Oleg Kambur Kipu Geneettisillä tekijöillä suuri merkitys Yksittäisiä geenejä on löydetty vain vähän COMT 1 Katekoli-O-metyylitransferaasi (COMT) proteiini tuotetaan
Oksidatiivinen fosforylaatio = ATP:n tuotto NADH:lta ja FADH2:lta hapelle tapahtuvan elektroninsiirron ja ATP-syntaasin avulla
 Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien elektronien vastaanottajana
Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien elektronien vastaanottajana
Ribosomit 1. Ribosomit 4. Ribosomit 2. Ribosomit 3. Proteiinisynteesin periaate 1
 Ribosomit 1 Ribosomit 4 Palade & Siekevitz eristivät jaottelusentrifugaatiolla ns. mikrosomeja radioakt. aminohapot kertyivät mikrosomeihin, jotka peräisin rer:ää sisältävistä soluista proteiinisynteesi
Ribosomit 1 Ribosomit 4 Palade & Siekevitz eristivät jaottelusentrifugaatiolla ns. mikrosomeja radioakt. aminohapot kertyivät mikrosomeihin, jotka peräisin rer:ää sisältävistä soluista proteiinisynteesi
Laskuharjoitus 2 vastauksia ja selityksiä
 Laskuharjoitus vastauksia ja selityksiä Selitykset ovat usein jonkin verran laajempia kuin vastaukseen edellytetyt, jotta asia selviäisi niillekin, jotka eivät sitä aluksi olleet keksineet. Tehtävä : Proteiinien
Laskuharjoitus vastauksia ja selityksiä Selitykset ovat usein jonkin verran laajempia kuin vastaukseen edellytetyt, jotta asia selviäisi niillekin, jotka eivät sitä aluksi olleet keksineet. Tehtävä : Proteiinien
sosiaaliturvatunnus Tehtävissä tarvittavia atomipainoja: hiili 12,01; vety 1,008; happi 16,00. Toisen asteen yhtälön ratkaisukaava: ax 2 + bx + c = 0;
 Valintakoe 2012 / Biokemia Nimi sosiaaliturvatunnus Tehtävissä tarvittavia atomipainoja: hiili 12,01; vety 1,008; happi 16,00. Toisen asteen yhtälön ratkaisukaava: ax 2 + bx + c = 0; 1. Kuvassa on esitetty
Valintakoe 2012 / Biokemia Nimi sosiaaliturvatunnus Tehtävissä tarvittavia atomipainoja: hiili 12,01; vety 1,008; happi 16,00. Toisen asteen yhtälön ratkaisukaava: ax 2 + bx + c = 0; 1. Kuvassa on esitetty
Syöpä. Ihmisen keho muodostuu miljardeista soluista. Vaikka. EGF-kasvutekijä. reseptori. tuma. dna
 Ihmisen keho muodostuu miljardeista soluista. Vaikka nämä solut ovat tietyssä mielessä meidän omiamme, ne polveutuvat itsenäisistä yksisoluisista elämänmuodoista, jotka ovat säilyttäneet monia itsenäisen
Ihmisen keho muodostuu miljardeista soluista. Vaikka nämä solut ovat tietyssä mielessä meidän omiamme, ne polveutuvat itsenäisistä yksisoluisista elämänmuodoista, jotka ovat säilyttäneet monia itsenäisen
Ionisidos ja ionihila:
 YHDISTEET KEMIAA KAIK- KIALLA, KE1 Ionisidos ja ionihila: Ionisidos syntyy kun metalli (pienempi elek.neg.) luovuttaa ulkoelektronin tai elektroneja epämetallille (elektronegatiivisempi). Ionisidos on
YHDISTEET KEMIAA KAIK- KIALLA, KE1 Ionisidos ja ionihila: Ionisidos syntyy kun metalli (pienempi elek.neg.) luovuttaa ulkoelektronin tai elektroneja epämetallille (elektronegatiivisempi). Ionisidos on
Siirtymämetallien erityisominaisuuksia
 Siirtymämetallien erityisominaisuuksia MATERIAALIT JA TEKNOLOGIA, KE4 Sivuryhmien metallien kemiaa: Jaksojen (vaakarivit) 4 ja 5 sivuryhmien metalleista käytetään myös nimitystä d-lohkon alkuaineet, koska
Siirtymämetallien erityisominaisuuksia MATERIAALIT JA TEKNOLOGIA, KE4 Sivuryhmien metallien kemiaa: Jaksojen (vaakarivit) 4 ja 5 sivuryhmien metalleista käytetään myös nimitystä d-lohkon alkuaineet, koska
Proteiinien muuntuminen energiaksi ihmiselimistössä
 Proteiinien muuntuminen energiaksi ihmiselimistössä Helsingin yliopisto Matemaattis-luonnontieteellinen tiedekunta Kemian laitos Kemian opettajankoulutusyksikkö Kandidaatintutkielma Tekijä: Klaus Sippel
Proteiinien muuntuminen energiaksi ihmiselimistössä Helsingin yliopisto Matemaattis-luonnontieteellinen tiedekunta Kemian laitos Kemian opettajankoulutusyksikkö Kandidaatintutkielma Tekijä: Klaus Sippel
Alikuoret eli orbitaalit
 Alkuaineiden jaksollinen järjestelmä Alkuaineen kemialliset ominaisuudet määräytyvät sen ulkokuoren elektronirakenteesta. Seuraus: Samanlaisen ulkokuorirakenteen omaavat alkuaineen ovat kemiallisesti sukulaisia
Alkuaineiden jaksollinen järjestelmä Alkuaineen kemialliset ominaisuudet määräytyvät sen ulkokuoren elektronirakenteesta. Seuraus: Samanlaisen ulkokuorirakenteen omaavat alkuaineen ovat kemiallisesti sukulaisia
? LUCA (Last universal common ancestor) 3.5 miljardia v.
 3.5 miljardia v.") Mitä elämä on? - Geneettinen ohjelma, joka kykenee muuttamaan ainehiukkaset ja molekyylit järjestyneeksi itseään replikoivaksi kokonaisuudeksi. (= geneettistä antientropiaa) ? LUCA (Last universal common
Mitä elämä on? - Geneettinen ohjelma, joka kykenee muuttamaan ainehiukkaset ja molekyylit järjestyneeksi itseään replikoivaksi kokonaisuudeksi. (= geneettistä antientropiaa) ? LUCA (Last universal common
CHEM-A1200 Kemiallinen rakenne ja sitoutuminen
 CHEM-A1200 Kemiallinen rakenne ja sitoutuminen Orgaaninen reaktio Opettava tutkija Pekka M Joensuu Orgaaniset reaktiot Syyt Pelkkä törmäys ei riitä Varaukset (myös osittaisvaraukset) houkuttelevat molekyylejä
CHEM-A1200 Kemiallinen rakenne ja sitoutuminen Orgaaninen reaktio Opettava tutkija Pekka M Joensuu Orgaaniset reaktiot Syyt Pelkkä törmäys ei riitä Varaukset (myös osittaisvaraukset) houkuttelevat molekyylejä
Vitamiinien puutostilat: Christian Ejkman (1858-1930) havaitsi ensimmäisenä vuonna 1888, että jonkin ravintotekijän puute aiheutti kanoilla
 havaitsi ensimmäisenä vuonna 1888, että jonkin ravintotekijän puute aiheutti kanoilla") Vitamiinit Vitamiinit ovat biologisesti aktiivisia orgaanisia yhdisteitä, joita elimistö tarvitsee, mutta ei itse pysty syntetisoimaan lainkaan tai riittävästi ja siksi niitä on saatava ravinnosta. Poikkeuksena
Vitamiinit Vitamiinit ovat biologisesti aktiivisia orgaanisia yhdisteitä, joita elimistö tarvitsee, mutta ei itse pysty syntetisoimaan lainkaan tai riittävästi ja siksi niitä on saatava ravinnosta. Poikkeuksena
Kaikenlaisia sidoksia yhdisteissä: ioni-, kovalenttiset ja metallisidokset Fysiikan ja kemian perusteet ja pedagogiikka
 Kaikenlaisia sidoksia yhdisteissä: ioni-, kovalenttiset ja metallisidokset Fysiikan ja kemian perusteet ja pedagogiikka Kari Sormunen Kevät 2012 Kertausta IONIEN MUODOSTUMISESTA Jos atomi luovuttaa tai
Kaikenlaisia sidoksia yhdisteissä: ioni-, kovalenttiset ja metallisidokset Fysiikan ja kemian perusteet ja pedagogiikka Kari Sormunen Kevät 2012 Kertausta IONIEN MUODOSTUMISESTA Jos atomi luovuttaa tai
BIOMOLEKYYLEJÄ. fruktoosi
 BIMLEKYYLEJÄ IMISEN JA ELINYMPÄ- RISTÖN KEMIAA, KE2 Ihminen on käyttänyt luonnosta saatavia, kasveissa ja eläimissä esiintyviä polymeerejä eli biopolymeerejä jo pitkään arkipäivän tarpeisiinsa. Biomolekyylit
BIMLEKYYLEJÄ IMISEN JA ELINYMPÄ- RISTÖN KEMIAA, KE2 Ihminen on käyttänyt luonnosta saatavia, kasveissa ja eläimissä esiintyviä polymeerejä eli biopolymeerejä jo pitkään arkipäivän tarpeisiinsa. Biomolekyylit
Etunimi: Henkilötunnus:
 Kokonaispisteet: Lue oheinen artikkeli ja vastaa kysymyksiin 1-25. Huomaa, että artikkelista ei löydy suoraan vastausta kaikkiin kysymyksiin, vaan sinun tulee myös tuntea ja selittää tarkemmin artikkelissa
Kokonaispisteet: Lue oheinen artikkeli ja vastaa kysymyksiin 1-25. Huomaa, että artikkelista ei löydy suoraan vastausta kaikkiin kysymyksiin, vaan sinun tulee myös tuntea ja selittää tarkemmin artikkelissa
Jonne Seppälä. Lectio praecursoria
 Jonne Seppälä Lectio praecursoria 22.5.2015 Structural Studies on Filamin Domain Interactions Rakennetutkimuksia filamiini-proteiinin domeenivuorovaikutuksilla Mitä solu- ja molekyylibioginen tutkimus
Jonne Seppälä Lectio praecursoria 22.5.2015 Structural Studies on Filamin Domain Interactions Rakennetutkimuksia filamiini-proteiinin domeenivuorovaikutuksilla Mitä solu- ja molekyylibioginen tutkimus
KE2 Kemian mikromaailma
 KE2 Kemian mikromaailma 30. maaliskuuta 2017/S.H. Vastaa viiteen tehtävään. Käytä tarvittaessa apuna taulukkokirjaa. Kopioi vastauspaperisi ensimmäisen sivun ylälaitaan seuraava taulukko. Kokeen pisteet
KE2 Kemian mikromaailma 30. maaliskuuta 2017/S.H. Vastaa viiteen tehtävään. Käytä tarvittaessa apuna taulukkokirjaa. Kopioi vastauspaperisi ensimmäisen sivun ylälaitaan seuraava taulukko. Kokeen pisteet
 Hoitotehoa ennustavat RAS-merkkiaineet Tärkeä apuväline kolorektaalisyövän lääkehoidon valinnassa Tämän esitteen tarkoitus Tämä esite auttaa ymmärtämään paremmin kolorektaalisyövän erilaisia lääkehoitovaihtoehtoja.
Hoitotehoa ennustavat RAS-merkkiaineet Tärkeä apuväline kolorektaalisyövän lääkehoidon valinnassa Tämän esitteen tarkoitus Tämä esite auttaa ymmärtämään paremmin kolorektaalisyövän erilaisia lääkehoitovaihtoehtoja.
Sytosoli eli solulima. Sytosoli. Solunsisäiset rakenteet, kalvostot ja proteiinien lajittelu (Chapter 12 Alberts et al.)
") Solunsisäiset rakenteet, kalvostot ja proteiinien lajittelu (Chapter 12 Alberts et al.) Figure 12-1 Molecular Biology of the Cell ( Garland Science 2008) Sytosoli eli solulima Sytosoli määritellään operatiivisesti
Solunsisäiset rakenteet, kalvostot ja proteiinien lajittelu (Chapter 12 Alberts et al.) Figure 12-1 Molecular Biology of the Cell ( Garland Science 2008) Sytosoli eli solulima Sytosoli määritellään operatiivisesti
2. Alkaanit. Suoraketjuiset alkaanit: etuliite+aani Metaani, etaani... Dekaani (10), undekaani, dodekaani, tridekaani, tetradekaani, pentadekaani..
, undekaani, dodekaani, tridekaani, tetradekaani, pentadekaani..") 2. Alkaanit SM -08 Kaikkein yksinkertaisimpia orgaanisia yhdisteitä. Sisältävät vain hiiltä ja vetyä ja vain yksinkertaisia - sidoksia. Yleinen molekyylikaava n 2n+2 Alkaanit voivat olla suoraketjuisia
2. Alkaanit SM -08 Kaikkein yksinkertaisimpia orgaanisia yhdisteitä. Sisältävät vain hiiltä ja vetyä ja vain yksinkertaisia - sidoksia. Yleinen molekyylikaava n 2n+2 Alkaanit voivat olla suoraketjuisia
MUUTOKSET ELEKTRONI- RAKENTEESSA
 MUUTOKSET ELEKTRONI- RAKENTEESSA KEMIAA KAIK- KIALLA, KE1 Ulkoelektronit ja oktettisääntö Alkuaineen korkeimmalla energiatasolla olevia elektroneja sanotaan ulkoelektroneiksi eli valenssielektroneiksi.
MUUTOKSET ELEKTRONI- RAKENTEESSA KEMIAA KAIK- KIALLA, KE1 Ulkoelektronit ja oktettisääntö Alkuaineen korkeimmalla energiatasolla olevia elektroneja sanotaan ulkoelektroneiksi eli valenssielektroneiksi.
Proteiinilääkkeet luento
 Proteiinilääkkeet luento 9.10.2017 DNA Bioteknologisen lääketuotannon periaate proteiini RNA RNA DNA proteiini Solu voi olla bakteeri, eläinsolu, tai tuotantoeläimen solu Ø Geneettisesti muokattu solulinja
Proteiinilääkkeet luento 9.10.2017 DNA Bioteknologisen lääketuotannon periaate proteiini RNA RNA DNA proteiini Solu voi olla bakteeri, eläinsolu, tai tuotantoeläimen solu Ø Geneettisesti muokattu solulinja
Isomerian lajit. Rakenne- eli konstituutioisomeria. Avaruus- eli stereoisomeria. Ketjuisomeria Funktioisomeria Paikkaisomeria
 Isomeria Isomeria Yhdisteellä on sama molekyylikaava, mutta eri rakenne: siksi eri isomeereillä voi olla erilaiset fysikaaliset ja kemialliset ominaisuudet!!!! Esim. yhdisteellä C2H6O on kaksi isomeeriä.
Isomeria Isomeria Yhdisteellä on sama molekyylikaava, mutta eri rakenne: siksi eri isomeereillä voi olla erilaiset fysikaaliset ja kemialliset ominaisuudet!!!! Esim. yhdisteellä C2H6O on kaksi isomeeriä.
BI4 IHMISEN BIOLOGIA
 BI4 IHMISEN BIOLOGIA 5 HORMONIT OVAT ELIMISTÖN TOIMINTAA SÄÄTELEVIÄ VIESTIAINEITA Avainsanat aivolisäke hormoni hypotalamus kasvuhormoni kortisoli palautesäätely rasvaliukoinen hormoni reseptori stressi
BI4 IHMISEN BIOLOGIA 5 HORMONIT OVAT ELIMISTÖN TOIMINTAA SÄÄTELEVIÄ VIESTIAINEITA Avainsanat aivolisäke hormoni hypotalamus kasvuhormoni kortisoli palautesäätely rasvaliukoinen hormoni reseptori stressi
Oulun yliopiston biokemian koulutusohjelman valintakoe 22.5.2015
 Oulun yliopiston biokemian koulutusohjelman valintakoe 22.5.2015 Nimi: Ota henkilötodistus mukaasi jättäessäsi vastauspaperin. Kysymyksiin vastataan suomeksi. Osa 1 Aineistotehtävä. Vastaa vain varattuun
Oulun yliopiston biokemian koulutusohjelman valintakoe 22.5.2015 Nimi: Ota henkilötodistus mukaasi jättäessäsi vastauspaperin. Kysymyksiin vastataan suomeksi. Osa 1 Aineistotehtävä. Vastaa vain varattuun
EPIONEN Kemia 2015. EPIONEN Kemia 2015
 EPIONEN Kemia 2015 1 Epione Valmennus 2014. Ensimmäinen painos www.epione.fi ISBN 978-952-5723-40-3 Painopaikka: Kopijyvä Oy, Kuopio Tämän teoksen painamiseen käytetty paperi on saanut Pohjoismaisen ympäristömerkin.
EPIONEN Kemia 2015 1 Epione Valmennus 2014. Ensimmäinen painos www.epione.fi ISBN 978-952-5723-40-3 Painopaikka: Kopijyvä Oy, Kuopio Tämän teoksen painamiseen käytetty paperi on saanut Pohjoismaisen ympäristömerkin.
Selluloosan rakenne ja ominaisuudet
 TEHTÄVÄ 1 - Pohjatiedot Selluloosan rakenne ja ominaisuudet 1. Millainen on selluloosan rakenne? 2. Missä selluloosa esiintyy soluseinässä? 3. Mikä on selluloosan tehtävä soluseinässä? Puu-19.210 Puun
TEHTÄVÄ 1 - Pohjatiedot Selluloosan rakenne ja ominaisuudet 1. Millainen on selluloosan rakenne? 2. Missä selluloosa esiintyy soluseinässä? 3. Mikä on selluloosan tehtävä soluseinässä? Puu-19.210 Puun
Solun perusrakenne I Solun perusrakenne. BI2 I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja
 Solun perusrakenne I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja 1. Avainsanat 2. Solut tuottavat entsyymejä katalyyteiksi 3. Entsyymien rakenne ja toiminta 4. Entsyymit vaativat toimiakseen
Solun perusrakenne I Solun perusrakenne 4. Entsyymit ovat solun kemiallisia robotteja 1. Avainsanat 2. Solut tuottavat entsyymejä katalyyteiksi 3. Entsyymien rakenne ja toiminta 4. Entsyymit vaativat toimiakseen
Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Tuomas Haltia Elämälle (solulle) välttämättömiä asioita ovat:
 välttämättömiä asioita ovat:") Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Tuomas Haltia 3.12.2012 Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien
Soluhengitys + ATP-synteesi = Oksidatiivinen fosforylaatio Tuomas Haltia 3.12.2012 Soluhengitys = Mitokondrioissa tapahtuva (ATP:tä tuottava) prosessi, jossa happi toimii pelkistyneiden ravintomolekyylien
- Jakautuvat kahteen selvästi erottuvaan luokkaan,
 Syöpä, osa II Syöpäkriittiset geenit - Geenejä, joiden mutaatiot usein havaitaan syöpien kanssa korreloituneena - Jakautuvat kahteen selvästi erottuvaan luokkaan, - dominoiviin onkogeeneihin - resessiivisiin
Syöpä, osa II Syöpäkriittiset geenit - Geenejä, joiden mutaatiot usein havaitaan syöpien kanssa korreloituneena - Jakautuvat kahteen selvästi erottuvaan luokkaan, - dominoiviin onkogeeneihin - resessiivisiin
d) Klooria valmistetaan hapettamalla vetykloridia kaliumpermanganaatilla. (Syntyy Mn 2+ -ioneja)
 Klooria valmistetaan hapettamalla vetykloridia kaliumpermanganaatilla. (Syntyy Mn 2+ -ioneja)") Helsingin yliopiston kemian valintakoe: Mallivastaukset. Maanantaina 29.5.2017 klo 14-17 1 Avogadron vakio NA = 6,022 10 23 mol -1 Yleinen kaasuvakio R = 8,314 J mol -1 K -1 = 0,08314 bar dm 3 mol -1 K
Helsingin yliopiston kemian valintakoe: Mallivastaukset. Maanantaina 29.5.2017 klo 14-17 1 Avogadron vakio NA = 6,022 10 23 mol -1 Yleinen kaasuvakio R = 8,314 J mol -1 K -1 = 0,08314 bar dm 3 mol -1 K
x _ Miksi elinikä ei ole rajaton? Mediterranean fruitfly (Ceratitis capitata) Eliniän jakautuma
 Eliniän jakautuma") Vanhenemisen fysiologia 1 Elinikä ja vanheneminen Käsitteet Elinikä - yksilön elinikä Eliniän odotusarvo, aktuaarinen (kirjanpidollinen) elinikä - keskimääräinen odotettavissa oleva elinikä tietylle lajille,
Vanhenemisen fysiologia 1 Elinikä ja vanheneminen Käsitteet Elinikä - yksilön elinikä Eliniän odotusarvo, aktuaarinen (kirjanpidollinen) elinikä - keskimääräinen odotettavissa oleva elinikä tietylle lajille,
Clinical impact of serum proteins on drug delivery Felix Kratz, Bakheet Elsadek Journal of Controlled Release 161 (2012)
") Clinical impact of serum proteins on drug delivery Felix Kratz, Bakheet Elsadek Journal of Controlled Release 161 (2012) 429 445 Sampo Kurvonen 25.10.2017 Sisältö Plasmaproteiineista Albumiini Transferriini
Clinical impact of serum proteins on drug delivery Felix Kratz, Bakheet Elsadek Journal of Controlled Release 161 (2012) 429 445 Sampo Kurvonen 25.10.2017 Sisältö Plasmaproteiineista Albumiini Transferriini
"Geenin toiminnan säätely" Moniste sivu 13
 "Geenin toiminnan säätely" Moniste sivu 13 Monisteen alussa on erittäin tärkeitä ohjeita turvallisuudesta Lukekaa sivu 5 huolellisesti ja usein Vaarat vaanivat: Palavia nesteitä ja liekkejä on joskus/usein
"Geenin toiminnan säätely" Moniste sivu 13 Monisteen alussa on erittäin tärkeitä ohjeita turvallisuudesta Lukekaa sivu 5 huolellisesti ja usein Vaarat vaanivat: Palavia nesteitä ja liekkejä on joskus/usein
Laskuharjoitus 3 palautus 11. 11. 2003 mennessä. Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa):
:") Laskuharjoitus 3 palautus 11. 11. 2003 mennessä Tehtävä 1: Entsyymikinetiikkaa Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa): 1. A:n sitoutuminen saa konformaatiossa aikaan muutoksen,
Laskuharjoitus 3 palautus 11. 11. 2003 mennessä Tehtävä 1: Entsyymikinetiikkaa Entsyymillä on seuraavanlainen reaktiomekanismi (katso oheista kuvaa): 1. A:n sitoutuminen saa konformaatiossa aikaan muutoksen,
class I T (Munz, autophagy (Argiris, 2008) 30 5 (Jemal, 2009) autophagy HLA / 4 21 (Sakakura, 2007; Chikamatsu, 2008; Chikamatsu, 2009) in vitro
 30 5 (Jemal, 2009) autophagy HLA / 4 21 (Sakakura, 2007; Chikamatsu, 2008; Chikamatsu, 2009) in vitro") 65 35 (Argiris, 2008)30 5 (Jemal, 2009) / 1991Boon / 4 21 (Sakakura, 2007; Chikamatsu, 2008; Chikamatsu, 2009) / / (Sakakura, 2005; Sakakura, 2006; Sakakura, 2007; Chikamatsu, 2007; Chikamatsu, 2008)/
65 35 (Argiris, 2008)30 5 (Jemal, 2009) / 1991Boon / 4 21 (Sakakura, 2007; Chikamatsu, 2008; Chikamatsu, 2009) / / (Sakakura, 2005; Sakakura, 2006; Sakakura, 2007; Chikamatsu, 2007; Chikamatsu, 2008)/
Biomolekyylit ja aineenvaihdunta I
 Biomolekyylit ja aineenvaihdunta I Luentorunko Jarmo Niemi 2003-2007 1 Biomolekyylit ja aineenvaihdunta I Luennot 6 op. (3 ov) 2003-2007 Jarmo Niemi (jarmo.niemi@utu.fi) Biokemian ja elintarvikekemian
Biomolekyylit ja aineenvaihdunta I Luentorunko Jarmo Niemi 2003-2007 1 Biomolekyylit ja aineenvaihdunta I Luennot 6 op. (3 ov) 2003-2007 Jarmo Niemi (jarmo.niemi@utu.fi) Biokemian ja elintarvikekemian
Designing switchable nanosystems for medical applica6on
 Designing switchable nanosystems for medical applica6on Lehner Roman, Wang Xueya, Wolf Marc, Hunziker Patrick Journal of controlled release 161:307-316, 2012 Emilia Karhunen 25.10.2017 22.11.2017 Emilia
Designing switchable nanosystems for medical applica6on Lehner Roman, Wang Xueya, Wolf Marc, Hunziker Patrick Journal of controlled release 161:307-316, 2012 Emilia Karhunen 25.10.2017 22.11.2017 Emilia
Geenisakset (CRISPR)- Geeniterapian vallankumousko? BMOL Juha Partanen
- Geeniterapian vallankumousko? BMOL Juha Partanen") Geenisakset (CRISPR)- Geeniterapian vallankumousko? BMOL 19.11.2016 Juha Partanen Geenisakset 2 2 N A T U R E V O L 5 2 2 4 J U N E 2 0 1 5 Sisältö Geenimuokkaus: historiallinen perspektiivi Geenisakset
Geenisakset (CRISPR)- Geeniterapian vallankumousko? BMOL 19.11.2016 Juha Partanen Geenisakset 2 2 N A T U R E V O L 5 2 2 4 J U N E 2 0 1 5 Sisältö Geenimuokkaus: historiallinen perspektiivi Geenisakset
8. Alkoholit, fenolit ja eetterit
 8. Alkoholit, fenolit ja eetterit SM -08 Alkoholit ovat orgaanisia yhdisteitä, joissa on yksi tai useampia -ryhmiä. Fenoleissa -ryhmä on kiinnittynyt aromaattiseen renkaaseen. Alkoholit voivat olla primäärisiä,
8. Alkoholit, fenolit ja eetterit SM -08 Alkoholit ovat orgaanisia yhdisteitä, joissa on yksi tai useampia -ryhmiä. Fenoleissa -ryhmä on kiinnittynyt aromaattiseen renkaaseen. Alkoholit voivat olla primäärisiä,
epiteeli endodermi Nisäkkään hampaan kehitys nisäkkään alkio:
 -mesenkyymi-vuorovaikutukset, esimerkkinä hammas ja ihokarva elimiä muodostuu kaikista alkiokerroksista, usein epiteelin ja mesenkyymin vuorovaikutuksesta epiteeli ektodermi kumpi aloittaa elimen kehityksen:
-mesenkyymi-vuorovaikutukset, esimerkkinä hammas ja ihokarva elimiä muodostuu kaikista alkiokerroksista, usein epiteelin ja mesenkyymin vuorovaikutuksesta epiteeli ektodermi kumpi aloittaa elimen kehityksen: