Regulatory Sequence Analysis. Discovering phylogenetic footprints in bacterial promoters
|
|
|
- Esa Kähkönen
- 6 vuotta sitten
- Katselukertoja:
Transkriptio
1 Regulatory Sequence Analysis Discovering phylogenetic footprints in bacterial promoters

2 DNA-protein binding interface Example of DNA-protein interface. LexA homodimer on the reca promoter. Note: this is only a model, the structure has still not been crystallized. Source: 2
3 LexA binding sites (RegulonDB, oct 2006) ECK reca b gaagcaattatactgtatgctcatacagtatcaagtgttt! ECK lexa b aaatcgccttttgctgtatatactcacagcataactgtat! ECK lexa b atactcacagcataactgtatatacacccagggggcggaa! ECK reca b gaagcaattatactgtatgctcatacagtatcaagtgttt! ECK ssb b gacacaaattgacctgaatgaatatacagtattggaatgc! ECK sula b agcccctgtgagttactgtatggatgtacagtacatccag! ECK uvra b gacacaaattgacctgaatgaatatacagtattggaatgc! ECK uvrb b ttatggtgatgaactgtttttttatccagtataatttgtt! ECK uvrd b taatcagcaaatctgtatatatacccagctttttggcgga! ECK rpsu b attttgaaataagctggcgttgatgccagcggcaaaccga! ECK phr b ttatcctgacgcctggctttcagggcagcgttatttcgaa! 3

4 LexA aligned binding sites and position-specific scoring matrix (PSSM) 1 1 : 1/7 ATTATACTGTATGCTCATACAGT! 2 2 : 2/8 CTTTTGCTGTATATACTCACAGC! 3 3 : 3/10 GCATAACTGTATATACACCCAGG! 4 4 : 4/7 ATTATACTGTATGCTCATACAGT! 5 5 : -5/13 CCAATACTGTATATTCATTCAGG! 6 6 : -6/15 GATGTACTGTACATCCATACAGT! 7 7 : -7/13 CCAATACTGTATATTCATTCAGG! 8 8 : -8/13 ATTATACTGGATAAAAAAACAGT! 9 9 : 9/7 GCAAATCTGTATATATACCCAGC! : 10/8 AATAAGCTGGCGTTGATGCCAGC! : -11/12 ATAACGCTGCCCTGAAAGCCAGG! Pos ! ! A ! C ! G ! T ! The site sequences were obtained from RegulonDB. The alignment and matrix were created using consensus (Hertz & Stormo, 1999). The logo was created with Web Logo ( 4
5 Analysis of Regulatory Sequences Phylogenetic footprint discovery in promoters of orthologous genes
6 Orthologs of Escherichia coli K12 gene PUTA in Gammaproteobacteria Orthologs of Escherichia coli K12 gene PUTA in Gammaproteobacteria NP_ Pasteurella_multocida!NP_ !52.93!0.0! YP_ Pseudoalteromonas_haloplanktis_TAC125!NP_ !45.21!0.0! YP_ Colwellia_psychrerythraea_34H!NP_ !43.81!0.0! NP_ Shewanella_oneidensis!NP_ !47.35!0.0! YP_ Idiomarina_loihiensis_L2TR!NP_ !45.44!0.0! YP_ Legionella_pneumophila_Philadelphia_1!NP_ !51.25!0.0! YP_ Legionella_pneumophila_Lens!NP_ !51.63!0.0! YP_ Legionella_pneumophila_Paris!NP_ !51.25!0.0! NP_ Coxiella_burnetii!NP_ !47.21!0.0! YP_ Photobacterium_profundum_SS9!NP_ !46.63!0.0! YP_ Vibrio_fischeri_ES114!NP_ !48.96!0.0! NP_ Vibrio_vulnificus_YJ016!NP_ !47.40!0.0! NP_ Vibrio_vulnificus_CMCP6!NP_ !47.60!0.0! NP_ Vibrio_parahaemolyticus!NP_ !47.92!0.0! NP_ Xanthomonas_campestris!NP_ !52.40!0.0! YP_ Xanthomonas_oryzae_KACC10331!NP_ !52.69!0.0! YP_ Xanthomonas_campestris_8004!NP_ !52.40!0.0! NP_ Xanthomonas_citri!NP_ !52.59!0.0! YP_ Xanthomonas_campestris_vesicatoria!NP_ !52.40!0.0! NP_ Salmonella_typhi_Ty2!NP_ !91.29!0.0! NP_ Salmonella_typhi!NP_ !91.29!0.0! YP_ Erwinia_carotovora_atrosepticaa!NP_ !74.24!0.0! YP_ Shigella_boydii_Sb227!NP_ !99.09!0.0! YP_ Shigella_sonnei_Ss046!NP_ !99.39!0.0! NP_ Escherichia_coli_CFT073!NP_ !99.39!0.0! NP_ Escherichia_coli_K12!NP_ !100.00!0.0! NP_ Escherichia_coli_O157H7!NP_ !99.55!0.0! NP_ Escherichia_coli_O157H7_EDL933!NP_ !99.55!0.0! NP_ Wigglesworthia_brevipalpis!NP_ !54.57!0.0! NP_ Yersinia_pestis_biovar_Mediaevails!NP_ !79.44!0.0! NP_ Yersinia_pestis_KIM!NP_ !79.37!0.0! YP_ Yersinia_pseudotuberculosis_IP32953!NP_ !79.37!0.0! NP_ Yersinia_pestis_CO92!NP_ !79.37!0.0! NP_ Photorhabdus_luminescens!NP_ !77.37!0.0! YP_ Nitrosococcus_oceani_ATCC_19707!NP_ !50.00!0.0! YP_ Francisella_tularensis_tularensis!NP_ !50.34!0.0! YP_ Hahella_chejuensis_KCTC_2396!NP_ !48.61!0.0! NP_ Pseudomonas_syringae!NP_ !73.64!0.0! NP_ Pseudomonas_putida_KT2440!NP_ !73.45!0.0! YP_ Pseudomonas_fluorescens_Pf-5!NP_ !73.60!0.0! YP_ Pseudomonas_fluorescens_PfO-1!NP_ !73.45!0.0! NP_ Pseudomonas_aeruginosa!NP_ !48.56!0.0! YP_ Pseudomonas_syringae_phaseolicola!NP_ !73.77!0.0! YP_ Pseudomonas_syringae_pv_B728a!NP_ !73.80!0.0! YP_ Psychrobacter_arcticum_273-4!NP_ !44.42!0.0! YP_ Acinetobacter_sp_ADP1!NP_ !59.51!0.0! 6
7 Upstream sequences of PUTA orthologs >YP_ Acinetobacter_sp_ADP1 puta!yp_ ; upstream from -103 to -1; size: 103;! ACAAAATTTTCCTCTAAAAAATGAATCAATTATAGTCAGTTATTTGGTAATTATTCTGTA! ATGATAAATTATCTAAACCCCTTTAAACAATATACCTTAGAGT! >YP_ Colwellia_psychrerythraea_34H puta!yp_ ; upstream from -245 to -1; size: 245! ATAATAACCCACGAACACTCCCTACAAATTATAAAAAACGATTTGCAGCACTTTATACTG! TTGAATTCGTACTCCCCCATATAAAAGTGTTTTAAATCCTGAAAATAACCAGCACATCCT! GTGGTTGTTACCCTAAAATTCGCTCATAAATTTAATGTCGTCACCAACTAATAATATATG! TATTAGTGGAAAAAAAGACTATAACTAAAGCAGGATTTCTACCTGTCACACTTTGAGGAA! TGGTT! >NP_ Coxiella_burnetii puta!np_ ; upstream from -118 to -1; size: 118;! GAAGTAGCCCGTATGAAGCGAAGCGAAATACGGGGAGGTGCACGTATTGTTCCCGTATTC! GCTTCGTTTCATACGGGCTACATCGCGGAAATGAAAATTAACTCCTTAATGAGGACAT! >YP_ Erwinia_carotovora_atroseptica_SCRI1043 puta!yp_ ; upstream from -195 to -1; size: 195;! TTAACTCTCCACATTTTTTTCGTGCCGCCGTCGCGGCGACGCTGTGTTTTATAGTAATCA! TTCAGGCCGGAACGAGGTCTGAAAATGATTATGGGCAGCAACCATTCCATTTGTTAACAA! GGTTGCACAAAGTTGCAACATGATTGATATTGACGGTATCCCGATGTGCATCTTCTCATT! ACAGGAGTGGACTCT! >NP_ Escherichia_coli_CFT073 puta!np_ ; upstream from 0 to -1; size: 0;! >NP_ Escherichia_coli_K12 puta!np_ ; upstream from -400 to -1; size: 400;! ATCGGCAATGTCGAAACTTGCCGTTATATCTGCCACCGGAACGGGGTAACAGAGTTTATG! TTTTACCAGGGCGACCGTATCCTGCCGGAAGCGCTGGTTATTCACAATCGATTTAACACA! CCATTTACATTAAATTTTAGTGCTCAGCGACACTATTTTTCATCAGGTTGCACTCTCTCA! CATTTTTTGCGGTTGCACCTTTCAAAAATGTTAACTGCCGCAGAGAAAAAGTCTGAGTTA! TTTTTTTAATCCCTGTCATATCGATTTCTTTTATTAACATTTCATTCATTTTTAAGCTTG! CTACGCATGTCACATTTAACATGGTTGCACAAAGTTGCAACATCATGGATATTTCACGAT! AACGTTAAGTTGCACCTTTCTGAACAACAGGAGTAATGGC! >NP_ Escherichia_coli_O157H7 ECs1260!NP_ ; upstream from -54 to -1; size: 54;! GGATATTTCACGATAACGTTAAGTTGCACCTTTCAGAACAACAGGAGTAATGGC! >NP_ Escherichia_coli_O157H7_EDL933 puta!np_ ; upstream from -400 to -1; size: 400;! ATCGGCAATGTCGAAACTTGCCGTTATATCTGCCACCGGAACGGGGTAACAGAGTTTATG! TTTTACCAGGGCGACCGTATCCTGCCGGAAGCGCTGGTTATTCACAATCGATTTAACACA! CCATTTACATTAAATTTTAGTGCTCAGCGACACTATTTTTCATCAGGTTGCACTCTCTCA! CATTTTTTGCGGTTGCACCTTTCAAAAATGTTAACTGCCGCAGAGAAAAAGTCTGAGTTA! TTTTTTTAATCCCTGTCATATCGATTTCTTTTATTAACATTTCATTCATTTTTAAGCTTG! CTACGCAGGTCACATTTAACATGGTTGCACAAAGTTGCAACATCATGGATATTTCACGAT! AACGTTAAGTTGCACCTTTCAGAACAACAGGAGTAATGGC!...! 7
8 Purged upstream sequences of PUTA orthologs >YP_ Acinetobacter_sp_ADP1 puta!! ACAAAATTTTCCTCTAAAAAATGAATCAATTATAGTCAGTTATTTGGTAATTATTCTGTA! ATGATAAATTATCTAAACCCCTTTAAACAATATACCTTAGAGT! >YP_ Colwellia_psychrerythraea_34H puta!! ATAATAACCCACGAACACTCCCTACAAATTATAAAAAACGATTTGCAGCACTTTATACTG! TTGAATTCGTACTCCCCCATATAAAAGTGTTTTAAATCCTGAAAATAACCAGCACATCCT! GTGGTTGTTACCCTAAAATTCGCTCATAAATTTAATGTCGTCACCAACTAATAATATATG! TATTAGTGGAAAAAAAGACTATAACTAAAGCAGGATTTCTACCTGTCACACTTTGAGGAA! TGGTT! >NP_ Coxiella_burnetii puta!! GAAGTAGCCCGTATGAAGCGAAGCGAAATACGGGGAGGTGCACGTATTGTTnnnnnnnnn! nnnnnnnnnnnnnnnnnnnnnatcgcggaaatgaaaattaactccttaatgaggacat! >YP_ Erwinia_carotovora_atroseptica_SCRI1043 puta!! TTAACTCTCCACATTTTTTTCGTGCCGCCGTCGCGGCGACGCTGTGTTTTATAGTAATCA! TTCAGGCCGGAACGAGGTCTGAAAATGATTATGGGCAGCAACCATTCCATTTGTTAACAA! GGTTGCACAAAGTTGCAACATGATTGATATTGACGGTATCCCGATGTGCATCTTCTCATT! ACAGGAGTGGACTCT! >NP_ Escherichia_coli_CFT073 puta!! >NP_ Escherichia_coli_K12 puta!! ATCGGCAATGTCGAAACTTGCCGTTATATCTGCCACCGGAACGGGGTAACAGAGTTTATG! TTTTACCAGGGCGACCGTATCCTGCCGGAAGCGCTGGTTATTCACAATCGATTTAACACA! CCATTTACATTAAATTTTAGTGCTCAGCGACACTATTTTTCATCAGGTTGCACTCTCTCA! CATTTTTTGCGGTTGCACCTTTCAAAAATGTTAACTGCCGCAGAGAAAAAGTCTGAGTTA! TTTTTTTAATCCCTGTCATATCGATTTCTTTTATTAACATTTCATTCATTTTTAAGCTTG! CTACGCATGTCACATnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnTCACGAT! AACGTTAAGTTGCACCTTTCTGAACAACAGGAGTAATGGC! >NP_ Escherichia_coli_O157H7 ECs1260!! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! >NP_ Escherichia_coli_O157H7_EDL933 puta!! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn! Nnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnn!...! 8
9 Significantly over-represented dyads in promoters of PUTA orthologs ; column headers ; 1 sequence ; 2 identifier ; 3 expected_freq ; 4 occ observed occurrences ; 5 exp_occ expected occurrences ; 6 occ_p occurrence probability (binomial) ; 7 occ_e E-value for occurrences (binomial) ; 8 occ_sig occurrence significance (binomial) ; 9 ovl_occ number of overlapping occurrences ; 10 all_occ number of non-overlapping + overlapping occurrences ; 11 rank rank ; 12 ov_coef overlap coefficient ; 13 remark remark ;sequence identifier expected_freocc exp_occ occ_p occ_e occ_sig ovl_occ all_occ rank ov_coef remark gtgn{1}aac gtgn{1}aac gttn{1}cac E E ggtn{3}acc ggtn{3}acc ggtn{3}acc E E inv_rep gttn{0}gca gttn{0}gca tgcn{0}aac E E ggtn{2}aac ggtn{2}aac gttn{2}acc E E gtgn{0}caa gtgn{0}caa ttgn{0}cac E E ggtn{2}cac ggtn{2}cac gtgn{2}acc E E ggtn{0}gca ggtn{0}gca tgcn{0}acc E E gcan{0}acc gcan{0}acc ggtn{0}tgc E E ggtn{1}caa ggtn{1}caa ttgn{1}acc E E ggtn{1}gca ggtn{1}gca tgcn{1}acc E E aagn{2}gca aagn{2}gca tgcn{2}ctt E E aggn{1}gca aggn{1}gca tgcn{1}cct E E aggn{0}tgc aggn{0}tgc gcan{0}cct E E aggn{2}caa aggn{2}caa ttgn{2}cct E E aagn{1}tgc aagn{1}tgc gcan{1}ctt E E aggn{3}aac aggn{3}aac gttn{3}cct E E gggn{4}acc gggn{4}acc ggtn{4}ccc E E ;Job started 13/02/06 21:31:01 CET ;Job done 13/02/06 21:31:28 CET 9
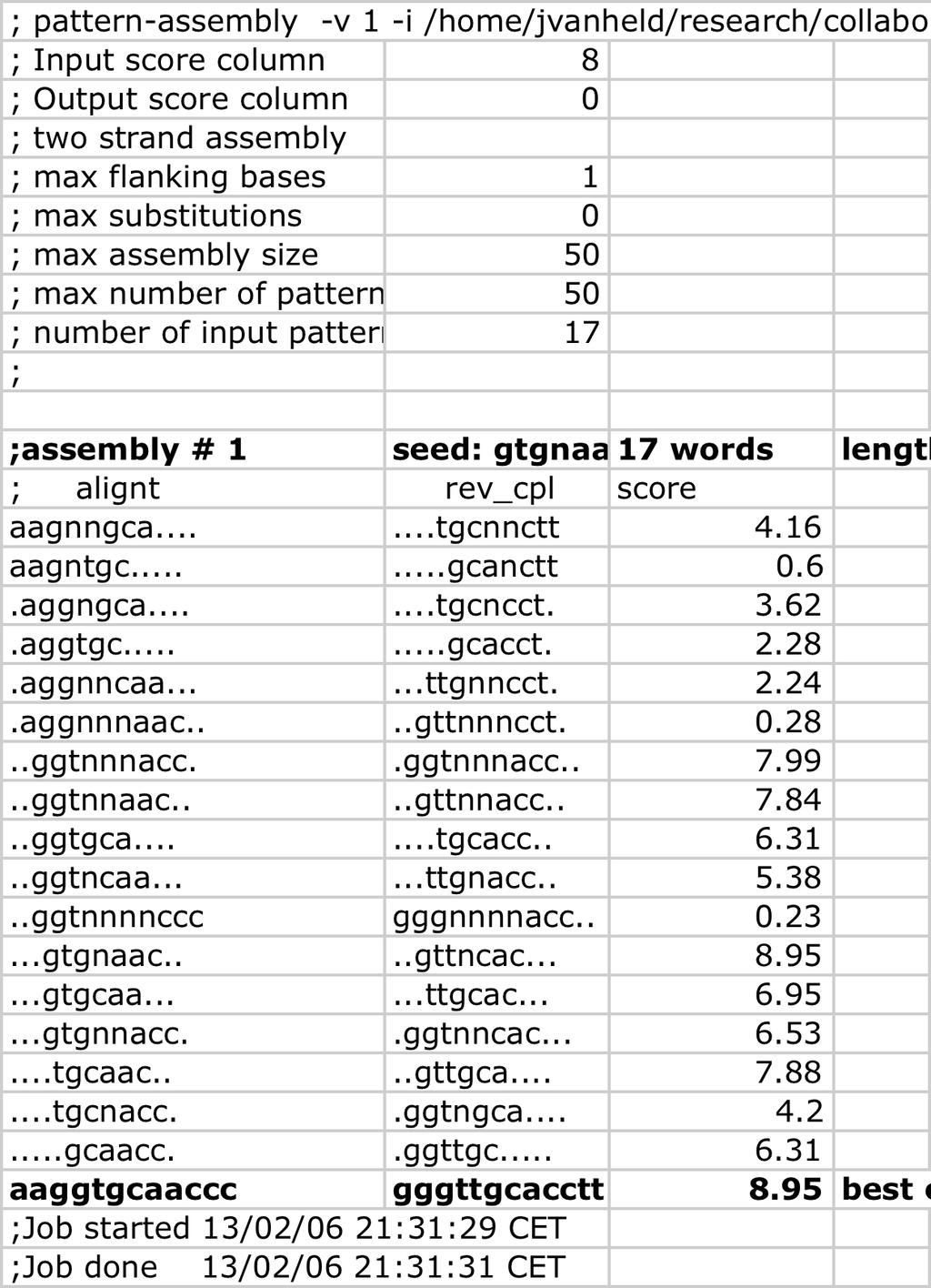

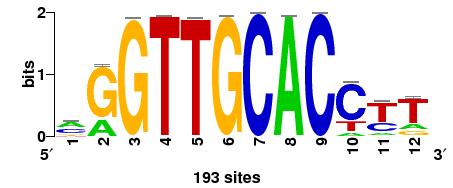
10 Significantly over-represented dyads in promoters of PUTA orthologs 10
11 Questions For each gene, we applied the same pattern discovery approach Identify orthologs Retrieve upstream sequences Detect over-represented dyads Questions How good is this method in predicting cis-acting elements? Can we detect correct motifs? What is the rate of false positives? Can we learn something about the evolution of cis-acting elements? On the basis of the discovered motifs, can we regroup the co-regulated genes? Detect pair-wise associations between genes Detect clusters of genes regulated by the same transcription factor 11
12 Analysis of Regulatory Sequences Evolution of the auto-regulation of the LexA transcription factor
13 Study case: LexA auto-regulation The transcription factor LexA represses several genes involved in the SOS response The lexa gene is auto-regulated. LexA auto-regulation has been characterized in details in several bacterial species. LexA protein has evolved and recognizes different motifs in different taxa. Note that this is an easy case: LexA binding motif is highly conserved in Gammaproteobacteria This motif is easily detected by most pattern discovery algorithms. We will start by this example, and then generalize the evaluation to other transcription factors. 13


.")
14 Evolution of lexa binding motifs Motifs characterized experimentally in different bacterial taxa Taxonomic level Species Consensus Weblogo Gamma-proteobacteria E.coli CTGTNNNNNNNNACAG Xanthomonadales Alpha proteobacteria X.oryzae X.campestris X.citri Xylella fastidiosa R.etli R.meliloti A.tumefasciens Rhodob acter sphaeroides Sinorhizob ium meliloti TTAGTArwawTACTAa GTTCNNNNNNNGTTC GAACNNNNNNNGAAC Myxococcus xanthus CTRHAMRYBYGTTCAGS Delta proteobacteria Geobacter sulfurreducens GGTTNNCNNNNGNNNACC Cyanobacteria Anabaena PCC7120 TAGTACTWATGTTCTA Gram+ bacteria B.sub tilis Mycob acterium tub erculosis Mycob acterium smegmatis CGAACRNRYGTTYC Janky, R. and van Helden, J. (2008). Evaluation of phylogenetic footprint discovery for predicting bacterial cis-regulatory elements and revealing their evolution. BMC Bioinformatics 9,

 Footprint-discovery result with lexa in")
15 Significant dyads in promoters of lexa orthologs in Gammaproteobacteria In Gammaproteobacteria, the most significant motif is aactgtatacatatacagtt This corresponds to the LexA motif annotated in RegulonDB Discovered motif (footprint-discovery) Annotated LexA binding motif (RegulonDB) Footprint-discovery result with lexa in Gammaproteobacteria, March

16 Significant dyads in promoters of lexa orthologs in Firmicutes In Firmicutes, the most significant motif is TACGAACATATGTTCGTA! This corresponds to the Cheo box GAACn 4 GTTC Janky, R. and van Helden, J. Evaluation of phylogenetic footprint discovery for predicting bacterial cis-regulatory elements and revealing their evolution BMC Bioinformatics 9, 37 (2008). 16
 In addition, this larger Cheo box is also detected in Actinobacteria, albeit with a")
17 Significant dyads in promoters of lexa orthologs in Actinobacteria In Actinobacteria, the most significant motif is TCGAACA. This shows an almost perfect match to one half of the Cheo box motif detected in Firmicutes (TACGAACATATGTTCGTA) In addition, this larger Cheo box is also detected in Actinobacteria, albeit with a lower significance than the half site. Janky, R. and van Helden, J. Evaluation of phylogenetic footprint discovery for predicting bacterial cis-regulatory elements and revealing their evolution BMC Bioinformatics 9, 37 (2008). 17









18 Significant dyads in promoters of lexa orthologs in Bacteria A Firmicutes Cyano. Actino. B C D When all the bacterial promoters are analyzed together, the program dyadanalysis detects most of taxon-specific motifs discussed before, and the feature-map highlights their taxon-specific locations. This illustrates the robustness of the method: the motifs can be detected even if present in a subset of the sequences only. Alpha. E The significance is however lower when all sequences are analyzed together than with the taxon-per-taxon analysis. Beta. F G H Gamma. Delta. Janky, R. and van Helden, J. Evaluation of phylogenetic footprint discovery for predicting bacterial cis-regulatory elements and revealing their evolution BMC Bioinformatics 9, 37 (2008). 18

19 Significance map of the motifs discovered in promoters of lexa orthologs at all taxonomical levels Gamma motif The heat map illustrates the most significant motifs (sig >= 8) found at different taxonomical levels. Each column corresponds to one dyad, each row to one taxon. The grey level indicates the significance. Firmicutes motif Xanthomonadales motif Alpha motif Beta motif Actinobacteria motif Janky, R. and van Helden, J. BMC Bioinformatics 9, 37 (2008). 19
20 Analysis of Regulatory Sequences Systematic evaluation of phylogenetic footprints discovery
21 Principle of the evaluation Choice of the sequence sets: Positive set: promoters of orthologs for all the genes having at least one annotated site in RegulonDB Negative set: random selections of promoters. Evaluation criteria Distribution of significance scores: does this score allow us to discriminate groups of co-regulated promoters from random selections? Motif correctness: do the motifs predicted in the positive set correspond to the annotated sites? 21
22 Evaluation of significance scores: distribution in promoters of orthologs versus random selections of genes Janky, R. and van Helden, J. BMC Bioinformatics 9, 37 (2008). 22
23 Correctness of the discovered motifs How to compare a discovered motif with a collection of annotated binding sites? The annotated binding sites can be considered as a set of oligonucleotides ( words ). The discovered motif is a set of dyads. We can compare two sets of words with the program compare-patterns. 23
24 Matching list - discovered dyads against annotated sites in glpd promoters Annotated sites for the gene glpd (Escherichia coli K12) Site sequence Factor site ID! gataaacgccataatgttatacatatcactctaaaatgtttt CRP ECK ! tctttgctaatatgttcgataacgaacatttatgagcttt GlpR ECK ! taacgaacatttatgagctttaacgaaagtgaatgagggc GlpR ECK ! gggatcaactggtttgcgttttggcgcaaattcagtgtta GlpR ECK ! aaaccggaaattaagcgcggattcgaatattctgactgtt GlpR ECK ! Perfect matches between detected dyads and annotated sites Site ID matching bases strand offset weight seq1 seq2 CRP_glpD_ECK R 30 6 AAAACATTTTAGAGTGATATGTATAACATTATGGCGTTTATC aacn{0}att GlpR_glpD_ECK D 11 6 tctttgctaatatgttcgataacgaacatttatgagcttt atgn{1}tcg GlpR_glpD_ECK D 24 6 tctttgctaatatgttcgataacgaacatttatgagcttt aacn{0}att GlpR_glpD_ECK D 22 6 tctttgctaatatgttcgataacgaacatttatgagcttt cgan{0}aca GlpR_glpD_ECK D 20 6 tctttgctaatatgttcgataacgaacatttatgagcttt aacn{2}aca GlpR_glpD_ECK R 20 6 AAAGCTCATAAATGTTCGTTATCGAACATATTAGCAAAGA aatn{2}tcg GlpR_glpD_ECK D 11 6 tctttgctaatatgttcgataacgaacatttatgagcttt atgn{0}ttc GlpR_glpD_ECK D 21 6 tctttgctaatatgttcgataacgaacatttatgagcttt acgn{2}cat GlpR_glpD_ECK D 20 6 tctttgctaatatgttcgataacgaacatttatgagcttt aacn{3}cat GlpR_glpD_ECK D 20 6 tctttgctaatatgttcgataacgaacatttatgagcttt aacn{0}gaa GlpR_glpD_ECK R 0 6 GCCCTCATTCACTTTCGTTAAAGCTCATAAATGTTCGTTA atgn{1}tcg GlpR_glpD_ECK D 5 6 taacgaacatttatgagctttaacgaaagtgaatgagggc aacn{0}att GlpR_glpD_ECK D 3 6 taacgaacatttatgagctttaacgaaagtgaatgagggc cgan{0}aca GlpR_glpD_ECK D 6 6 taacgaacatttatgagctttaacgaaagtgaatgagggc acan{13}acg GlpR_glpD_ECK D 1 6 taacgaacatttatgagctttaacgaaagtgaatgagggc aacn{2}aca GlpR_glpD_ECK R 1 6 GCCCTCATTCACTTTCGTTAAAGCTCATAAATGTTCGTTA aatn{2}tcg GlpR_glpD_ECK R 0 6 GCCCTCATTCACTTTCGTTAAAGCTCATAAATGTTCGTTA atgn{0}ttc GlpR_glpD_ECK D 2 6 taacgaacatttatgagctttaacgaaagtgaatgagggc acgn{2}cat GlpR_glpD_ECK D 1 6 taacgaacatttatgagctttaacgaaagtgaatgagggc aacn{3}cat GlpR_glpD_ECK R 28 6 GCCCTCATTCACTTTCGTTAAAGCTCATAAATGTTCGTTA cctn{4}cac GlpR_glpD_ECK D 1 6 taacgaacatttatgagctttaacgaaagtgaatgagggc aacn{0}gaa GlpR_glpD_ECK R 20 6 GCCCTCATTCACTTTCGTTAAAGCTCATAAATGTTCGTTA actn{7}aag GlpR_glpD_ECK R 21 6 AACAGTCAGAATATTCGAATCCGCGCTTAATTTCCGGTTT aatn{2}tcg GlpR_glpD_ECK D 1 6 aaaccggaaattaagcgcggattcgaatattctgactgtt aacn{8}aag 24
25 Matching table - discovered dyads against annotated sites in glpd promoters The dyad-site cross-table allows us to define correspondence statistics. Sensitivity (Sn) Sn = TP/(TP+FN) fraction of annotated sites matched by at least one discovered dyad Positive Predictive Value (PPV) PPV = TP/(TP+FP) Fraction of discovered dyads matching at least one annotated site Accuracy (geometric) Acc=sqrt(Sn PPV) Geometric mean of Sn and PPV gataaacgccataatgttatacatatcactctaaaatgtttt tctttgctaatatgttcgataacgaacatttatgagcttt taacgaacatttatgagctttaacgaaagtgaatgagggc gggatcaactggtttgcgttttggcgcaaattcagtgtta aaaccggaaattaagcgcggattcgaatattctgactgtt ; sequence atgn{1}tcg aacn{0}att cgan{0}aca agtn{10}tcg..... acan{13}acg aacn{2}aca aatn{2}tcg atgn{0}ttc acgn{2}cat aacn{3}cat aacn{8}aag cctn{4}cac aacn{0}gaa actn{7}aag
26 Correctness statistics for all the genes having at least one annotated site in RegulonDB 26
27 Supplementary material
28 Discovered motifs in promoters of glpd orthologs from Enterobacteriales Dyad exp_freq occ exp_occ occ_p occ_e sig rank atgn{1}tcg cgan{1}cat E E aacn{0}att aatn{0}gtt E E cgan{0}aca tgtn{0}tcg E E agtn{10}tcg cgan{10}act E E acan{13}acg cgtn{13}tgt E E aacn{2}aca tgtn{2}gtt E E aatn{2}tcg cgan{2}att E E atgn{0}ttc gaan{0}cat E E acgn{2}cat atgn{2}cgt E E aacn{3}cat atgn{3}gtt E E aacn{8}aag cttn{8}gtt E E cctn{4}cac gtgn{4}agg E E aacn{0}gaa ttcn{0}gtt E E actn{7}aag cttn{7}agt E E ;assembly # 1 seed: atgntcg 13 words aaangtt......aacnttt 0.82.aatgtt......aacatt aatnntcg....cgannatt atgntcg....cgancat atgttc......gaacat atgnnngtt aacnnncat atgnncgt..acgnncat tgttcg....cgaaca tgtnngtt aacnnaca tgtncgt..acgnaca gttngtt aacnaac gttcgt..acgaac ttcgtt aacgaa aaatgttcgtt aacgaacattt 7.55 ;assembly # 2 seed: atgnnnnnnntcg 19 words aaangtt......aacnttt 0.82.aatgtt......aacatt aatnntcg......cgannatt atgntcg......cgancat atgttc......gaacat atgnnnnnnntcg..cgannnnnnncat atgnnngtt......aacnnncat atgnncgt......acgnncat atgnnnnnnnncga tcgnnnnnnnncat tgttcg......cgaaca tgtnngtt......aacnnaca tgtncgt......acgnaca tgtnnnnnntcg..cgannnnnnaca gttngtt......aacnaac gttcgt......acgaac gttnnnnntcg..cgannnnnaac ttcgtt......aacgaa tcgnnntcg..cgannncga tcgnnnncga tcgnnnncga aaatgttcgttntcga tcganaacgaacattt 7.55 ;assembly # 3 seed: agtnnnnnnnnnntcg 3 words agtnnnnnnnnnntcg cgannnnnnnnnnact 1.52.gttnnnnnnnnntcg cgannnnnnnnnaac ttcnnnnnnnntcg cgannnnnnnngaa agttcnnnnnnnntcg cgannnnnnnngaact 1.52 ;assembly # 4 seed: acannnnnnnnnnnnnacg 3 words acannnnnnnnnnnnnacg..cgtnnnnnnnnnnnnntgt 1.26 acannnnnnnnnnnnnncga tcgnnnnnnnnnnnnnntgt 0.48 acannnnnnnnnnnnaac....gttnnnnnnnnnnnntgt 0.01 acannnnnnnnnnnnaacga tcgttnnnnnnnnnnnntgt
Chapter 7. Motif finding (week 11) Chapter 8. Sequence binning (week 11)
 Chapter 8. Sequence binning (week 11)") Course organization Introduction ( Week 1) Part I: Algorithms for Sequence Analysis (Week 1-11) Chapter 1-3, Models and theories» Probability theory and Statistics (Week 2)» Algorithm complexity analysis
Course organization Introduction ( Week 1) Part I: Algorithms for Sequence Analysis (Week 1-11) Chapter 1-3, Models and theories» Probability theory and Statistics (Week 2)» Algorithm complexity analysis
Efficiency change over time
 Efficiency change over time Heikki Tikanmäki Optimointiopin seminaari 14.11.2007 Contents Introduction (11.1) Window analysis (11.2) Example, application, analysis Malmquist index (11.3) Dealing with panel
Efficiency change over time Heikki Tikanmäki Optimointiopin seminaari 14.11.2007 Contents Introduction (11.1) Window analysis (11.2) Example, application, analysis Malmquist index (11.3) Dealing with panel
Other approaches to restrict multipliers
 Other approaches to restrict multipliers Heikki Tikanmäki Optimointiopin seminaari 10.10.2007 Contents Short revision (6.2) Another Assurance Region Model (6.3) Cone-Ratio Method (6.4) An Application of
Other approaches to restrict multipliers Heikki Tikanmäki Optimointiopin seminaari 10.10.2007 Contents Short revision (6.2) Another Assurance Region Model (6.3) Cone-Ratio Method (6.4) An Application of
Alternative DEA Models
 Mat-2.4142 Alternative DEA Models 19.9.2007 Table of Contents Banker-Charnes-Cooper Model Additive Model Example Data Home assignment BCC Model (Banker-Charnes-Cooper) production frontiers spanned by convex
Mat-2.4142 Alternative DEA Models 19.9.2007 Table of Contents Banker-Charnes-Cooper Model Additive Model Example Data Home assignment BCC Model (Banker-Charnes-Cooper) production frontiers spanned by convex
FETAL FIBROBLASTS, PASSAGE 10
 Double-stranded methylation patterns of a 104-bp L1 promoter in DNAs from fetal fibroblast passages 10, 14, 17, and 22 using barcoded hairpinbisulfite PCR. Fifteen L1 sequences were analyzed for passages
Double-stranded methylation patterns of a 104-bp L1 promoter in DNAs from fetal fibroblast passages 10, 14, 17, and 22 using barcoded hairpinbisulfite PCR. Fifteen L1 sequences were analyzed for passages
Bounds on non-surjective cellular automata
 Bounds on non-surjective cellular automata Jarkko Kari Pascal Vanier Thomas Zeume University of Turku LIF Marseille Universität Hannover 27 august 2009 J. Kari, P. Vanier, T. Zeume (UTU) Bounds on non-surjective
Bounds on non-surjective cellular automata Jarkko Kari Pascal Vanier Thomas Zeume University of Turku LIF Marseille Universität Hannover 27 august 2009 J. Kari, P. Vanier, T. Zeume (UTU) Bounds on non-surjective
Gap-filling methods for CH 4 data
 Gap-filling methods for CH 4 data Sigrid Dengel University of Helsinki Outline - Ecosystems known for CH 4 emissions; - Why is gap-filling of CH 4 data not as easy and straight forward as CO 2 ; - Gap-filling
Gap-filling methods for CH 4 data Sigrid Dengel University of Helsinki Outline - Ecosystems known for CH 4 emissions; - Why is gap-filling of CH 4 data not as easy and straight forward as CO 2 ; - Gap-filling
MALE ADULT FIBROBLAST LINE (82-6hTERT)
") Double-stranded methylation patterns of a 104-bp L1 promoter in DNAs from male and female fibroblasts, male leukocytes and female lymphoblastoid cells using hairpin-bisulfite PCR. Fifteen L1 sequences
Double-stranded methylation patterns of a 104-bp L1 promoter in DNAs from male and female fibroblasts, male leukocytes and female lymphoblastoid cells using hairpin-bisulfite PCR. Fifteen L1 sequences
On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31)
") On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31) Juha Kahkonen Click here if your download doesn"t start automatically On instrument costs
On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31) Juha Kahkonen Click here if your download doesn"t start automatically On instrument costs
11/17/11. Gene Regulation. Gene Regulation. Gene Regulation. Finding Regulatory Motifs in DNA Sequences. Regulatory Proteins
 Gene Regulation Finding Regulatory Motifs in DNA Sequences An experiment shows that when X is knocked out, 20 other s are not expressed How can one have such drastic effects? Regulatory Proteins Gene X
Gene Regulation Finding Regulatory Motifs in DNA Sequences An experiment shows that when X is knocked out, 20 other s are not expressed How can one have such drastic effects? Regulatory Proteins Gene X
Basset: Learning the regulatory code of the accessible genome with deep convolutional neural networks. David R. Kelley
 Basset: Learning the regulatory code of the accessible genome with deep convolutional neural networks David R. Kelley DNA codes for complex life. How? Kundaje et al. Integrative analysis of 111 reference
Basset: Learning the regulatory code of the accessible genome with deep convolutional neural networks David R. Kelley DNA codes for complex life. How? Kundaje et al. Integrative analysis of 111 reference
Eukaryotic Comparative Genomics
 Eukaryotic Comparative Genomics Detecting Conserved Sequences Charles Darwin Motoo Kimura Evolution of Neutral DNA A A T C TA AT T G CT G T GA T T C A GA G T A G CA G T GA AT A GT C T T T GA T GT T G T
Eukaryotic Comparative Genomics Detecting Conserved Sequences Charles Darwin Motoo Kimura Evolution of Neutral DNA A A T C TA AT T G CT G T GA T T C A GA G T A G CA G T GA AT A GT C T T T GA T GT T G T
tgg agg Supplementary Figure S1.
 ttaggatattcggtgaggtgatatgtctctgtttggaaatgtctccgccattaactcaag tggaaagtgtatagtaatgaatctttcaagcacacagatcacttcaaaagactgtttcaa catcacctcaggacaaaaagatgtactctcatttggatgctgtgatgccatgggtcacag attgcaattcccaagtgcccgttcttttacaccaaaatcaaagaagaatatctccccttt
ttaggatattcggtgaggtgatatgtctctgtttggaaatgtctccgccattaactcaag tggaaagtgtatagtaatgaatctttcaagcacacagatcacttcaaaagactgtttcaa catcacctcaggacaaaaagatgtactctcatttggatgctgtgatgccatgggtcacag attgcaattcccaagtgcccgttcttttacaccaaaatcaaagaagaatatctccccttt
Returns to Scale II. S ysteemianalyysin. Laboratorio. Esitelmä 8 Timo Salminen. Teknillinen korkeakoulu
 Returns to Scale II Contents Most Productive Scale Size Further Considerations Relaxation of the Convexity Condition Useful Reminder Theorem 5.5 A DMU found to be efficient with a CCR model will also be
Returns to Scale II Contents Most Productive Scale Size Further Considerations Relaxation of the Convexity Condition Useful Reminder Theorem 5.5 A DMU found to be efficient with a CCR model will also be
Chapter 9 Motif finding. Chaochun Wei Spring 2019
 1896 1920 1987 2006 Chapter 9 Motif finding Chaochun Wei Spring 2019 Contents 1. Reading materials 2. Sequence structure modeling Motif finding Regulatory module finding 2 Reading materials Tompa et al
1896 1920 1987 2006 Chapter 9 Motif finding Chaochun Wei Spring 2019 Contents 1. Reading materials 2. Sequence structure modeling Motif finding Regulatory module finding 2 Reading materials Tompa et al
T Statistical Natural Language Processing Answers 6 Collocations Version 1.0
 T-61.5020 Statistical Natural Language Processing Answers 6 Collocations Version 1.0 1. Let s start by calculating the results for pair valkoinen, talo manually: Frequency: Bigrams valkoinen, talo occurred
T-61.5020 Statistical Natural Language Processing Answers 6 Collocations Version 1.0 1. Let s start by calculating the results for pair valkoinen, talo manually: Frequency: Bigrams valkoinen, talo occurred
Methods S1. Sequences relevant to the constructed strains, Related to Figures 1-6.
 Methods S1. Sequences relevant to the constructed strains, Related to Figures 1-6. A. Promoter Sequences Gal4 binding sites are highlighted in the color referenced in Figure 1A when possible. Site 1: red,
Methods S1. Sequences relevant to the constructed strains, Related to Figures 1-6. A. Promoter Sequences Gal4 binding sites are highlighted in the color referenced in Figure 1A when possible. Site 1: red,
The CCR Model and Production Correspondence
 The CCR Model and Production Correspondence Tim Schöneberg The 19th of September Agenda Introduction Definitions Production Possiblity Set CCR Model and the Dual Problem Input excesses and output shortfalls
The CCR Model and Production Correspondence Tim Schöneberg The 19th of September Agenda Introduction Definitions Production Possiblity Set CCR Model and the Dual Problem Input excesses and output shortfalls
Capacity Utilization
 Capacity Utilization Tim Schöneberg 28th November Agenda Introduction Fixed and variable input ressources Technical capacity utilization Price based capacity utilization measure Long run and short run
Capacity Utilization Tim Schöneberg 28th November Agenda Introduction Fixed and variable input ressources Technical capacity utilization Price based capacity utilization measure Long run and short run
Network to Get Work. Tehtäviä opiskelijoille Assignments for students. www.laurea.fi
 Network to Get Work Tehtäviä opiskelijoille Assignments for students www.laurea.fi Ohje henkilöstölle Instructions for Staff Seuraavassa on esitetty joukko tehtäviä, joista voit valita opiskelijaryhmällesi
Network to Get Work Tehtäviä opiskelijoille Assignments for students www.laurea.fi Ohje henkilöstölle Instructions for Staff Seuraavassa on esitetty joukko tehtäviä, joista voit valita opiskelijaryhmällesi
KONEISTUSKOKOONPANON TEKEMINEN NX10-YMPÄRISTÖSSÄ
 KONEISTUSKOKOONPANON TEKEMINEN NX10-YMPÄRISTÖSSÄ https://community.plm.automation.siemens.com/t5/tech-tips- Knowledge-Base-NX/How-to-simulate-any-G-code-file-in-NX- CAM/ta-p/3340 Koneistusympäristön määrittely
KONEISTUSKOKOONPANON TEKEMINEN NX10-YMPÄRISTÖSSÄ https://community.plm.automation.siemens.com/t5/tech-tips- Knowledge-Base-NX/How-to-simulate-any-G-code-file-in-NX- CAM/ta-p/3340 Koneistusympäristön määrittely
812336A C++ -kielen perusteet, 21.8.2010
 812336A C++ -kielen perusteet, 21.8.2010 1. Vastaa lyhyesti seuraaviin kysymyksiin (1p kaikista): a) Mitä tarkoittaa funktion ylikuormittaminen (overloading)? b) Mitä tarkoittaa jäsenfunktion ylimääritys
812336A C++ -kielen perusteet, 21.8.2010 1. Vastaa lyhyesti seuraaviin kysymyksiin (1p kaikista): a) Mitä tarkoittaa funktion ylikuormittaminen (overloading)? b) Mitä tarkoittaa jäsenfunktion ylimääritys
FinFamily PostgreSQL installation ( ) FinFamily PostgreSQL
 FinFamily PostgreSQL") FinFamily PostgreSQL 1 Sisällys / Contents FinFamily PostgreSQL... 1 1. Asenna PostgreSQL tietokanta / Install PostgreSQL database... 3 1.1. PostgreSQL tietokannasta / About the PostgreSQL database...
FinFamily PostgreSQL 1 Sisällys / Contents FinFamily PostgreSQL... 1 1. Asenna PostgreSQL tietokanta / Install PostgreSQL database... 3 1.1. PostgreSQL tietokannasta / About the PostgreSQL database...
Plasmid Name: pmm290. Aliases: none known. Length: bp. Constructed by: Mike Moser/Cristina Swanson. Last updated: 17 August 2009
 Plasmid Name: pmm290 Aliases: none known Length: 11707 bp Constructed by: Mike Moser/Cristina Swanson Last updated: 17 August 2009 Description and application: This is a mammalian expression vector for
Plasmid Name: pmm290 Aliases: none known Length: 11707 bp Constructed by: Mike Moser/Cristina Swanson Last updated: 17 August 2009 Description and application: This is a mammalian expression vector for
Statistical design. Tuomas Selander
 Statistical design Tuomas Selander 28.8.2014 Introduction Biostatistician Work area KYS-erva KYS, Jyväskylä, Joensuu, Mikkeli, Savonlinna Work tasks Statistical methods, selection and quiding Data analysis
Statistical design Tuomas Selander 28.8.2014 Introduction Biostatistician Work area KYS-erva KYS, Jyväskylä, Joensuu, Mikkeli, Savonlinna Work tasks Statistical methods, selection and quiding Data analysis
On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31)
") On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31) Juha Kahkonen Click here if your download doesn"t start automatically On instrument costs
On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31) Juha Kahkonen Click here if your download doesn"t start automatically On instrument costs
Searching (Sub-)Strings. Ulf Leser
Strings. Ulf Leser") Searching (Sub-)Strings Ulf Leser This Lecture Exact substring search Naïve Boyer-Moore Searching with profiles Sequence profiles Ungapped approximate search Statistical evaluation of search results Ulf
Searching (Sub-)Strings Ulf Leser This Lecture Exact substring search Naïve Boyer-Moore Searching with profiles Sequence profiles Ungapped approximate search Statistical evaluation of search results Ulf
Choose Finland-Helsinki Valitse Finland-Helsinki
 Write down the Temporary Application ID. If you do not manage to complete the form you can continue where you stopped with this ID no. Muista Temporary Application ID. Jos et onnistu täyttää lomake loppuun
Write down the Temporary Application ID. If you do not manage to complete the form you can continue where you stopped with this ID no. Muista Temporary Application ID. Jos et onnistu täyttää lomake loppuun
Results on the new polydrug use questions in the Finnish TDI data
 Results on the new polydrug use questions in the Finnish TDI data Multi-drug use, polydrug use and problematic polydrug use Martta Forsell, Finnish Focal Point 28/09/2015 Martta Forsell 1 28/09/2015 Esityksen
Results on the new polydrug use questions in the Finnish TDI data Multi-drug use, polydrug use and problematic polydrug use Martta Forsell, Finnish Focal Point 28/09/2015 Martta Forsell 1 28/09/2015 Esityksen
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
Functional Genomics & Proteomics
 Functional Genomics & Proteomics Genome Sequences TCACAATTTAGACATCTAGTCTTCCACTTAAGCATATTTAGATTGTTTCCAGTTTTCAGCTTTTATGACTAAATCTTCTAAAATTGTTTTTCCCTAAATGTATATTTTAATTTGTCTCAGGAGTAGAATTTCTGAGTCATAAAGCGGT CATATGTATAAATTTTAGGTGCCTCATAGCTCTTCAAATAGTCATCCCATTTTATACATCCAGGCAATATATGAGAGTTCTTGGTGCTCCACATCTTAGCTAGGATTTGATGTCAACCAGTCTCTTTAATTTAGATATTCTAGTACAT
Functional Genomics & Proteomics Genome Sequences TCACAATTTAGACATCTAGTCTTCCACTTAAGCATATTTAGATTGTTTCCAGTTTTCAGCTTTTATGACTAAATCTTCTAAAATTGTTTTTCCCTAAATGTATATTTTAATTTGTCTCAGGAGTAGAATTTCTGAGTCATAAAGCGGT CATATGTATAAATTTTAGGTGCCTCATAGCTCTTCAAATAGTCATCCCATTTTATACATCCAGGCAATATATGAGAGTTCTTGGTGCTCCACATCTTAGCTAGGATTTGATGTCAACCAGTCTCTTTAATTTAGATATTCTAGTACAT
Supporting information
 Supporting information Figure S1. Carotenoid biosynthesis pathway in papaya fruit which is adopted from Blas et al. (2010) (reference 4) and Nisar et al. (2015) (reference 5). Carotenoids are synthesized
Supporting information Figure S1. Carotenoid biosynthesis pathway in papaya fruit which is adopted from Blas et al. (2010) (reference 4) and Nisar et al. (2015) (reference 5). Carotenoids are synthesized
On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31)
") On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31) Juha Kahkonen Click here if your download doesn"t start automatically On instrument costs
On instrument costs in decentralized macroeconomic decision making (Helsingin Kauppakorkeakoulun julkaisuja ; D-31) Juha Kahkonen Click here if your download doesn"t start automatically On instrument costs
Metsälamminkankaan tuulivoimapuiston osayleiskaava
 VAALAN KUNTA TUULISAIMAA OY Metsälamminkankaan tuulivoimapuiston osayleiskaava Liite 3. Varjostusmallinnus FCG SUUNNITTELU JA TEKNIIKKA OY 12.5.2015 P25370 SHADOW - Main Result Assumptions for shadow calculations
VAALAN KUNTA TUULISAIMAA OY Metsälamminkankaan tuulivoimapuiston osayleiskaava Liite 3. Varjostusmallinnus FCG SUUNNITTELU JA TEKNIIKKA OY 12.5.2015 P25370 SHADOW - Main Result Assumptions for shadow calculations
make and make and make ThinkMath 2017
 Adding quantities Lukumäärienup yhdistäminen. Laske yhteensä?. Countkuinka howmonta manypalloja ballson there are altogether. and ja make and make and ja make on and ja make ThinkMath 7 on ja on on Vaihdannaisuus
Adding quantities Lukumäärienup yhdistäminen. Laske yhteensä?. Countkuinka howmonta manypalloja ballson there are altogether. and ja make and make and ja make on and ja make ThinkMath 7 on ja on on Vaihdannaisuus
Skene. Games Refueled. Muokkaa perustyyl. napsautt. @Games for Health, Kuopio. 2013 kari.korhonen@tekes.fi. www.tekes.fi/skene
 Skene Muokkaa perustyyl. Games Refueled napsautt. @Games for Health, Kuopio Muokkaa alaotsikon perustyyliä napsautt. 2013 kari.korhonen@tekes.fi www.tekes.fi/skene 10.9.201 3 Muokkaa Skene boosts perustyyl.
Skene Muokkaa perustyyl. Games Refueled napsautt. @Games for Health, Kuopio Muokkaa alaotsikon perustyyliä napsautt. 2013 kari.korhonen@tekes.fi www.tekes.fi/skene 10.9.201 3 Muokkaa Skene boosts perustyyl.
Paikkatiedon semanttinen mallinnus, integrointi ja julkaiseminen Case Suomalainen ajallinen paikkaontologia SAPO
 Paikkatiedon semanttinen mallinnus, integrointi ja julkaiseminen Case Suomalainen ajallinen paikkaontologia SAPO Tomi Kauppinen, Eero Hyvönen, Jari Väätäinen Semantic Computing Research Group (SeCo) http://www.seco.tkk.fi/
Paikkatiedon semanttinen mallinnus, integrointi ja julkaiseminen Case Suomalainen ajallinen paikkaontologia SAPO Tomi Kauppinen, Eero Hyvönen, Jari Väätäinen Semantic Computing Research Group (SeCo) http://www.seco.tkk.fi/
1. SIT. The handler and dog stop with the dog sitting at heel. When the dog is sitting, the handler cues the dog to heel forward.
 START START SIT 1. SIT. The handler and dog stop with the dog sitting at heel. When the dog is sitting, the handler cues the dog to heel forward. This is a static exercise. SIT STAND 2. SIT STAND. The
START START SIT 1. SIT. The handler and dog stop with the dog sitting at heel. When the dog is sitting, the handler cues the dog to heel forward. This is a static exercise. SIT STAND 2. SIT STAND. The
4x4cup Rastikuvien tulkinta
 4x4cup Rastikuvien tulkinta 4x4cup Control point picture guidelines Päivitetty kauden 2010 sääntöihin Updated for 2010 rules Säännöt rastikuvista Kilpailijoiden tulee kiinnittää erityistä huomiota siihen,
4x4cup Rastikuvien tulkinta 4x4cup Control point picture guidelines Päivitetty kauden 2010 sääntöihin Updated for 2010 rules Säännöt rastikuvista Kilpailijoiden tulee kiinnittää erityistä huomiota siihen,
Experimental Identification and Computational Characterization of a Novel. Extracellular Metalloproteinase Produced by Clostridium sordellii
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 207 Supplementary Information Experimental Identification and Computational Characterization of
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 207 Supplementary Information Experimental Identification and Computational Characterization of
C++11 seminaari, kevät Johannes Koskinen
 C++11 seminaari, kevät 2012 Johannes Koskinen Sisältö Mikä onkaan ongelma? Standardidraftin luku 29: Atomiset tyypit Muistimalli Rinnakkaisuus On multicore systems, when a thread writes a value to memory,
C++11 seminaari, kevät 2012 Johannes Koskinen Sisältö Mikä onkaan ongelma? Standardidraftin luku 29: Atomiset tyypit Muistimalli Rinnakkaisuus On multicore systems, when a thread writes a value to memory,
Use of spatial data in the new production environment and in a data warehouse
 Use of spatial data in the new production environment and in a data warehouse Nordic Forum for Geostatistics 2007 Session 3, GI infrastructure and use of spatial database Statistics Finland, Population
Use of spatial data in the new production environment and in a data warehouse Nordic Forum for Geostatistics 2007 Session 3, GI infrastructure and use of spatial database Statistics Finland, Population
( ( OX2 Perkkiö. Rakennuskanta. Varjostus. 9 x N131 x HH145
 OX2 9 x N131 x HH145 Rakennuskanta Asuinrakennus Lomarakennus Liike- tai julkinen rakennus Teollinen rakennus Kirkko tai kirkollinen rak. Muu rakennus Allas Varjostus 1 h/a 8 h/a 20 h/a 0 0,5 1 1,5 2 km
OX2 9 x N131 x HH145 Rakennuskanta Asuinrakennus Lomarakennus Liike- tai julkinen rakennus Teollinen rakennus Kirkko tai kirkollinen rak. Muu rakennus Allas Varjostus 1 h/a 8 h/a 20 h/a 0 0,5 1 1,5 2 km
16. Allocation Models
 16. Allocation Models Juha Saloheimo 17.1.27 S steemianalsin Optimointiopin seminaari - Sks 27 Content Introduction Overall Efficienc with common prices and costs Cost Efficienc S steemianalsin Revenue
16. Allocation Models Juha Saloheimo 17.1.27 S steemianalsin Optimointiopin seminaari - Sks 27 Content Introduction Overall Efficienc with common prices and costs Cost Efficienc S steemianalsin Revenue
Tynnyrivaara, OX2 Tuulivoimahanke. ( Layout 9 x N131 x HH145. Rakennukset Asuinrakennus Lomarakennus 9 x N131 x HH145 Varjostus 1 h/a 8 h/a 20 h/a
 , Tuulivoimahanke Layout 9 x N131 x HH145 Rakennukset Asuinrakennus Lomarakennus 9 x N131 x HH145 Varjostus 1 h/a 8 h/a 20 h/a 0 0,5 1 1,5 km 2 SHADOW - Main Result Assumptions for shadow calculations
, Tuulivoimahanke Layout 9 x N131 x HH145 Rakennukset Asuinrakennus Lomarakennus 9 x N131 x HH145 Varjostus 1 h/a 8 h/a 20 h/a 0 0,5 1 1,5 km 2 SHADOW - Main Result Assumptions for shadow calculations
Information on preparing Presentation
 Information on preparing Presentation Seminar on big data management Lecturer: Spring 2017 20.1.2017 1 Agenda Hints and tips on giving a good presentation Watch two videos and discussion 22.1.2017 2 Goals
Information on preparing Presentation Seminar on big data management Lecturer: Spring 2017 20.1.2017 1 Agenda Hints and tips on giving a good presentation Watch two videos and discussion 22.1.2017 2 Goals
Kysymys 5 Compared to the workload, the number of credits awarded was (1 credits equals 27 working hours): (4)
: (4)") Tilasto T1106120-s2012palaute Kyselyn T1106120+T1106120-s2012palaute yhteenveto: vastauksia (4) Kysymys 1 Degree programme: (4) TIK: TIK 1 25% ************** INF: INF 0 0% EST: EST 0 0% TLT: TLT 0 0% BIO:
Tilasto T1106120-s2012palaute Kyselyn T1106120+T1106120-s2012palaute yhteenveto: vastauksia (4) Kysymys 1 Degree programme: (4) TIK: TIK 1 25% ************** INF: INF 0 0% EST: EST 0 0% TLT: TLT 0 0% BIO:
WindPRO version joulu 2012 Printed/Page :42 / 1. SHADOW - Main Result
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table 13.6.2013 19:42 / 1 Minimum
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table 13.6.2013 19:42 / 1 Minimum
Valuation of Asian Quanto- Basket Options
 Valuation of Asian Quanto- Basket Options (Final Presentation) 21.11.2011 Thesis Instructor and Supervisor: Prof. Ahti Salo Työn saa tallentaa ja julkistaa Aalto-yliopiston avoimilla verkkosivuilla. Muilta
Valuation of Asian Quanto- Basket Options (Final Presentation) 21.11.2011 Thesis Instructor and Supervisor: Prof. Ahti Salo Työn saa tallentaa ja julkistaa Aalto-yliopiston avoimilla verkkosivuilla. Muilta
Telecommunication Software
 Telecommunication Software Final exam 21.11.2006 COMPUTER ENGINEERING LABORATORY 521265A Vastaukset englanniksi tai suomeksi. / Answers in English or in Finnish. 1. (a) Määrittele sovellusviesti, PersonnelRecord,
Telecommunication Software Final exam 21.11.2006 COMPUTER ENGINEERING LABORATORY 521265A Vastaukset englanniksi tai suomeksi. / Answers in English or in Finnish. 1. (a) Määrittele sovellusviesti, PersonnelRecord,
Predicting evolutionarily conserved regions (ECRs) in the Xenopus tropicalis genome using a MultiPipMaker-based bioinformatic strategy
 in the Xenopus tropicalis genome using a MultiPipMaker-based bioinformatic strategy") Predicting evolutionarily conserved regions (ECRs) in the Xenopus tropicalis genome using a MultiPipMaker-based bioinformatic strategy OVERVIEW- Method updated and written by Sarah Louie (6/19/08) Initially
Predicting evolutionarily conserved regions (ECRs) in the Xenopus tropicalis genome using a MultiPipMaker-based bioinformatic strategy OVERVIEW- Method updated and written by Sarah Louie (6/19/08) Initially
RINNAKKAINEN OHJELMOINTI A,
 RINNAKKAINEN OHJELMOINTI 815301A, 18.6.2005 1. Vastaa lyhyesti (2p kustakin): a) Mitkä ovat rinnakkaisen ohjelman oikeellisuuskriteerit? b) Mitä tarkoittaa laiska säikeen luominen? c) Mitä ovat kohtaaminen
RINNAKKAINEN OHJELMOINTI 815301A, 18.6.2005 1. Vastaa lyhyesti (2p kustakin): a) Mitkä ovat rinnakkaisen ohjelman oikeellisuuskriteerit? b) Mitä tarkoittaa laiska säikeen luominen? c) Mitä ovat kohtaaminen
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
LYTH-CONS CONSISTENCY TRANSMITTER
 LYTH-CONS CONSISTENCY TRANSMITTER LYTH-INSTRUMENT OY has generate new consistency transmitter with blade-system to meet high technical requirements in Pulp&Paper industries. Insurmountable advantages are
LYTH-CONS CONSISTENCY TRANSMITTER LYTH-INSTRUMENT OY has generate new consistency transmitter with blade-system to meet high technical requirements in Pulp&Paper industries. Insurmountable advantages are
( ,5 1 1,5 2 km
 Tuulivoimala Rakennukset Asuinrakennus Liikerak. tai Julkinen rak. Lomarakennus Teollinen rakennus Kirkollinen rakennus Varjostus "real case" h/a 1 h/a 8 h/a 20 h/a 4 5 3 1 2 6 7 8 9 10 0 0,5 1 1,5 2 km
Tuulivoimala Rakennukset Asuinrakennus Liikerak. tai Julkinen rak. Lomarakennus Teollinen rakennus Kirkollinen rakennus Varjostus "real case" h/a 1 h/a 8 h/a 20 h/a 4 5 3 1 2 6 7 8 9 10 0 0,5 1 1,5 2 km
The role of 3dr sector in rural -community based- tourism - potentials, challenges
 The role of 3dr sector in rural -community based- tourism - potentials, challenges Lappeenranta, 5th September 2014 Contents of the presentation 1. SEPRA what is it and why does it exist? 2. Experiences
The role of 3dr sector in rural -community based- tourism - potentials, challenges Lappeenranta, 5th September 2014 Contents of the presentation 1. SEPRA what is it and why does it exist? 2. Experiences
Uusi Ajatus Löytyy Luonnosta 4 (käsikirja) (Finnish Edition)
 (Finnish Edition)") Uusi Ajatus Löytyy Luonnosta 4 (käsikirja) (Finnish Edition) Esko Jalkanen Click here if your download doesn"t start automatically Uusi Ajatus Löytyy Luonnosta 4 (käsikirja) (Finnish Edition) Esko Jalkanen
Uusi Ajatus Löytyy Luonnosta 4 (käsikirja) (Finnish Edition) Esko Jalkanen Click here if your download doesn"t start automatically Uusi Ajatus Löytyy Luonnosta 4 (käsikirja) (Finnish Edition) Esko Jalkanen
WindPRO version joulu 2012 Printed/Page :47 / 1. SHADOW - Main Result
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table 22.12.2014 11:33 / 1 Minimum
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table 22.12.2014 11:33 / 1 Minimum
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Calculation: N117 x 9 x HH141 Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG
SHADOW - Main Result Calculation: N117 x 9 x HH141 Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG
GOOD WORK LONGER CAREER:
 Juhani Ilmarinen, Ville Ilmarinen, Pekka Huuhtanen, Veikko Louhevaara, Ove Näsman GOOD WORK LONGER CAREER: WORK WELL-BEING IN FINNISH TECHNOLOGY INDUSTRIES 2010-2015 Background Collective agreement between
Juhani Ilmarinen, Ville Ilmarinen, Pekka Huuhtanen, Veikko Louhevaara, Ove Näsman GOOD WORK LONGER CAREER: WORK WELL-BEING IN FINNISH TECHNOLOGY INDUSTRIES 2010-2015 Background Collective agreement between
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.9.269
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.9.269
Capacity utilization
 Mat-2.4142 Seminar on optimization Capacity utilization 12.12.2007 Contents Summary of chapter 14 Related DEA-solver models Illustrative examples Measure of technical capacity utilization Price-based measure
Mat-2.4142 Seminar on optimization Capacity utilization 12.12.2007 Contents Summary of chapter 14 Related DEA-solver models Illustrative examples Measure of technical capacity utilization Price-based measure
Digital Admap Native. Campaign: Kesko supermarket
 Digital Admap Native Campaign: Kesko supermarket Digital Admap Native Campaign: Kesko Supermarket Mainosmuoto: Natiivi Media: IS.fi Campaign period: 25 September Date of measurement: 26 September Unique:
Digital Admap Native Campaign: Kesko supermarket Digital Admap Native Campaign: Kesko Supermarket Mainosmuoto: Natiivi Media: IS.fi Campaign period: 25 September Date of measurement: 26 September Unique:
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
Salasanan vaihto uuteen / How to change password
 Salasanan vaihto uuteen / How to change password Sisällys Salasanakäytäntö / Password policy... 2 Salasanan vaihto verkkosivulla / Change password on website... 3 Salasanan vaihto matkapuhelimella / Change
Salasanan vaihto uuteen / How to change password Sisällys Salasanakäytäntö / Password policy... 2 Salasanan vaihto verkkosivulla / Change password on website... 3 Salasanan vaihto matkapuhelimella / Change
Infrastruktuurin asemoituminen kansalliseen ja kansainväliseen kenttään Outi Ala-Honkola Tiedeasiantuntija
 Infrastruktuurin asemoituminen kansalliseen ja kansainväliseen kenttään Outi Ala-Honkola Tiedeasiantuntija 1 Asemoitumisen kuvaus Hakemukset parantuneet viime vuodesta, mutta paneeli toivoi edelleen asemoitumisen
Infrastruktuurin asemoituminen kansalliseen ja kansainväliseen kenttään Outi Ala-Honkola Tiedeasiantuntija 1 Asemoitumisen kuvaus Hakemukset parantuneet viime vuodesta, mutta paneeli toivoi edelleen asemoitumisen
TM ETRS-TM35FIN-ETRS89 WTG
 VE1 SHADOW - Main Result Calculation: 8 x Nordex N131 x HH145m Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please
VE1 SHADOW - Main Result Calculation: 8 x Nordex N131 x HH145m Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please
1.3Lohkorakenne muodostetaan käyttämällä a) puolipistettä b) aaltosulkeita c) BEGIN ja END lausekkeita d) sisennystä
 puolipistettä b) aaltosulkeita c) BEGIN ja END lausekkeita d) sisennystä") OULUN YLIOPISTO Tietojenkäsittelytieteiden laitos Johdatus ohjelmointiin 81122P (4 ov.) 30.5.2005 Ohjelmointikieli on Java. Tentissä saa olla materiaali mukana. Tenttitulokset julkaistaan aikaisintaan
OULUN YLIOPISTO Tietojenkäsittelytieteiden laitos Johdatus ohjelmointiin 81122P (4 ov.) 30.5.2005 Ohjelmointikieli on Java. Tentissä saa olla materiaali mukana. Tenttitulokset julkaistaan aikaisintaan
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table 5.11.2013 16:44 / 1 Minimum
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table 5.11.2013 16:44 / 1 Minimum
SELL Student Games kansainvälinen opiskelijaurheilutapahtuma
 SELL Student Games kansainvälinen opiskelijaurheilutapahtuma Painonnosto 13.5.2016 (kansallinen, CUP) Below in English Paikka: Nääshalli Näsijärvenkatu 8 33210 Tampere Alustava aikataulu: Punnitus 12:00-13:00
SELL Student Games kansainvälinen opiskelijaurheilutapahtuma Painonnosto 13.5.2016 (kansallinen, CUP) Below in English Paikka: Nääshalli Näsijärvenkatu 8 33210 Tampere Alustava aikataulu: Punnitus 12:00-13:00
7.4 Variability management
 7.4 Variability management time... space software product-line should support variability in space (different products) support variability in time (maintenance, evolution) 1 Product variation Product
7.4 Variability management time... space software product-line should support variability in space (different products) support variability in time (maintenance, evolution) 1 Product variation Product
anna minun kertoa let me tell you
 anna minun kertoa let me tell you anna minun kertoa I OSA 1. Anna minun kertoa sinulle mitä oli. Tiedän että osaan. Kykenen siihen. Teen nyt niin. Minulla on oikeus. Sanani voivat olla puutteellisia mutta
anna minun kertoa let me tell you anna minun kertoa I OSA 1. Anna minun kertoa sinulle mitä oli. Tiedän että osaan. Kykenen siihen. Teen nyt niin. Minulla on oikeus. Sanani voivat olla puutteellisia mutta
State of the Union... Functional Genomics Research Stream. Molecular Biology. Genomics. Computational Biology
 Functional Genomics Research Stream State of the Union... Research Meeting: February 16, 2010 Functional Genomics & Research Report III Concepts Genomics Molecular Biology Computational Biology Genome
Functional Genomics Research Stream State of the Union... Research Meeting: February 16, 2010 Functional Genomics & Research Report III Concepts Genomics Molecular Biology Computational Biology Genome
,0 Yes ,0 120, ,8
 SHADOW - Main Result Calculation: Alue 2 ( x 9 x HH120) TuuliSaimaa kaavaluonnos Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered
SHADOW - Main Result Calculation: Alue 2 ( x 9 x HH120) TuuliSaimaa kaavaluonnos Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered
4x4cup Rastikuvien tulkinta. 4x4cup Control point picture guidelines
 4x4cup Rastikuvien tulkinta 4x4cup Control point picture guidelines Säännöt rastikuvista Kilpailijoiden tulee kiinnittää erityistä huomiota siihen, että rastikuvissa näkyy selvästi että kilpailija koskee
4x4cup Rastikuvien tulkinta 4x4cup Control point picture guidelines Säännöt rastikuvista Kilpailijoiden tulee kiinnittää erityistä huomiota siihen, että rastikuvissa näkyy selvästi että kilpailija koskee
RANTALA SARI: Sairaanhoitajan eettisten ohjeiden tunnettavuus ja niiden käyttö hoitotyön tukena sisätautien vuodeosastolla
 TURUN YLIOPISTO Hoitotieteen laitos RANTALA SARI: Sairaanhoitajan eettisten ohjeiden tunnettavuus ja niiden käyttö hoitotyön tukena sisätautien vuodeosastolla Pro gradu -tutkielma, 34 sivua, 10 liitesivua
TURUN YLIOPISTO Hoitotieteen laitos RANTALA SARI: Sairaanhoitajan eettisten ohjeiden tunnettavuus ja niiden käyttö hoitotyön tukena sisätautien vuodeosastolla Pro gradu -tutkielma, 34 sivua, 10 liitesivua
 Digitally signed by Hans Vadbäck DN: cn=hans Vadbäck, o, ou=fcg Suunnittelu ja Tekniikka Oy, email=hans.vadback@fcg.fi, c=fi Date: 2016.12.20 15:45:35 +02'00' Jakob Kjellman Digitally signed by Jakob Kjellman
Digitally signed by Hans Vadbäck DN: cn=hans Vadbäck, o, ou=fcg Suunnittelu ja Tekniikka Oy, email=hans.vadback@fcg.fi, c=fi Date: 2016.12.20 15:45:35 +02'00' Jakob Kjellman Digitally signed by Jakob Kjellman
Operatioanalyysi 2011, Harjoitus 2, viikko 38
 Operatioanalyysi 2011, Harjoitus 2, viikko 38 H2t1, Exercise 1.1. H2t2, Exercise 1.2. H2t3, Exercise 2.3. H2t4, Exercise 2.4. H2t5, Exercise 2.5. (Exercise 1.1.) 1 1.1. Model the following problem mathematically:
Operatioanalyysi 2011, Harjoitus 2, viikko 38 H2t1, Exercise 1.1. H2t2, Exercise 1.2. H2t3, Exercise 2.3. H2t4, Exercise 2.4. H2t5, Exercise 2.5. (Exercise 1.1.) 1 1.1. Model the following problem mathematically:
Information on Finnish Courses Autumn Semester 2017 Jenni Laine & Päivi Paukku Centre for Language and Communication Studies
 Information on Finnish Courses Autumn Semester 2017 Jenni Laine & Päivi Paukku 24.8.2017 Centre for Language and Communication Studies Puhutko suomea? -Hei! -Hei hei! -Moi! -Moi moi! -Terve! -Terve terve!
Information on Finnish Courses Autumn Semester 2017 Jenni Laine & Päivi Paukku 24.8.2017 Centre for Language and Communication Studies Puhutko suomea? -Hei! -Hei hei! -Moi! -Moi moi! -Terve! -Terve terve!
Information on Finnish Language Courses Spring Semester 2017 Jenni Laine
 Information on Finnish Language Courses Spring Semester 2017 Jenni Laine 4.1.2017 KIELIKESKUS LANGUAGE CENTRE Puhutko suomea? Do you speak Finnish? -Hei! -Moi! -Mitä kuuluu? -Kiitos, hyvää. -Entä sinulle?
Information on Finnish Language Courses Spring Semester 2017 Jenni Laine 4.1.2017 KIELIKESKUS LANGUAGE CENTRE Puhutko suomea? Do you speak Finnish? -Hei! -Moi! -Mitä kuuluu? -Kiitos, hyvää. -Entä sinulle?
Ajettavat luokat: SM: S1 (25 aika-ajon nopeinta)
") SUPERMOTO SM 2013 OULU Lisämääräys ja ohje Oulun Moottorikerho ry ja Oulun Formula K-125ry toivottaa SuperMoto kuljettajat osallistumaan SuperMoto SM 2013 Oulu osakilpailuun. Kilpailu ajetaan karting radalla
SUPERMOTO SM 2013 OULU Lisämääräys ja ohje Oulun Moottorikerho ry ja Oulun Formula K-125ry toivottaa SuperMoto kuljettajat osallistumaan SuperMoto SM 2013 Oulu osakilpailuun. Kilpailu ajetaan karting radalla
ELEMET- MOCASTRO. Effect of grain size on A 3 temperatures in C-Mn and low alloyed steels - Gleeble tests and predictions. Period
 1 ELEMET- MOCASTRO Effect of grain size on A 3 temperatures in C-Mn and low alloyed steels - Gleeble tests and predictions Period 20.02-25.05.2012 Diaarinumero Rahoituspäätöksen numero 1114/31/2010 502/10
1 ELEMET- MOCASTRO Effect of grain size on A 3 temperatures in C-Mn and low alloyed steels - Gleeble tests and predictions Period 20.02-25.05.2012 Diaarinumero Rahoituspäätöksen numero 1114/31/2010 502/10
A DEA Game II. Juha Saloheimo S ysteemianalyysin. Laboratorio. Teknillinen korkeakoulu
 A DEA Game II Juha Salohemo 12.12.2007 Content Recap of the Example The Shapley Value Margnal Contrbuton, Ordered Coaltons, Soluton to the Example DEA Mn Game Summary Home Assgnment Recap of the Example
A DEA Game II Juha Salohemo 12.12.2007 Content Recap of the Example The Shapley Value Margnal Contrbuton, Ordered Coaltons, Soluton to the Example DEA Mn Game Summary Home Assgnment Recap of the Example
Voice Over LTE (VoLTE) By Miikka Poikselkä;Harri Holma;Jukka Hongisto
 By Miikka Poikselkä;Harri Holma;Jukka Hongisto") Voice Over LTE (VoLTE) By Miikka Poikselkä;Harri Holma;Jukka Hongisto If you are searched for a book by Miikka Poikselkä;Harri Holma;Jukka Hongisto Voice over LTE (VoLTE) in pdf form, then you have come
Voice Over LTE (VoLTE) By Miikka Poikselkä;Harri Holma;Jukka Hongisto If you are searched for a book by Miikka Poikselkä;Harri Holma;Jukka Hongisto Voice over LTE (VoLTE) in pdf form, then you have come
TM ETRS-TM35FIN-ETRS89 WTG
 SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
SHADOW - Main Result Assumptions for shadow calculations Maximum distance for influence Calculate only when more than 20 % of sun is covered by the blade Please look in WTG table WindPRO version 2.8.579
Co-Design Yhteissuunnittelu
 Co-Design Yhteissuunnittelu Tuuli Mattelmäki DA, associate professor Aalto University School of Arts, Design and Architecture School of Arts, Design and Architecture design with and for people Codesign
Co-Design Yhteissuunnittelu Tuuli Mattelmäki DA, associate professor Aalto University School of Arts, Design and Architecture School of Arts, Design and Architecture design with and for people Codesign
VAASAN YLIOPISTO Humanististen tieteiden kandidaatin tutkinto / Filosofian maisterin tutkinto
 VAASAN YLIOPISTO Humanististen tieteiden kandidaatin tutkinto / Filosofian maisterin tutkinto Tämän viestinnän, nykysuomen ja englannin kandidaattiohjelman valintakokeen avulla Arvioidaan viestintävalmiuksia,
VAASAN YLIOPISTO Humanististen tieteiden kandidaatin tutkinto / Filosofian maisterin tutkinto Tämän viestinnän, nykysuomen ja englannin kandidaattiohjelman valintakokeen avulla Arvioidaan viestintävalmiuksia,
National Building Code of Finland, Part D1, Building Water Supply and Sewerage Systems, Regulations and guidelines 2007
 National Building Code of Finland, Part D1, Building Water Supply and Sewerage Systems, Regulations and guidelines 2007 Chapter 2.4 Jukka Räisä 1 WATER PIPES PLACEMENT 2.4.1 Regulation Water pipe and its
National Building Code of Finland, Part D1, Building Water Supply and Sewerage Systems, Regulations and guidelines 2007 Chapter 2.4 Jukka Räisä 1 WATER PIPES PLACEMENT 2.4.1 Regulation Water pipe and its
ReFuel 70 % Emission Reduction Using Renewable High Cetane Number Paraffinic Diesel Fuel. Kalle Lehto, Aalto-yliopisto 5.5.
 ReFuel 70 % Emission Reduction Using Renewable High Cetane Number Paraffinic Diesel Fuel Kalle Lehto, Aalto-yliopisto 5.5.2011 Otaniemi ReFuel a three year research project (2009-2011) goal utilize the
ReFuel 70 % Emission Reduction Using Renewable High Cetane Number Paraffinic Diesel Fuel Kalle Lehto, Aalto-yliopisto 5.5.2011 Otaniemi ReFuel a three year research project (2009-2011) goal utilize the
Information on Finnish Language Courses Spring Semester 2018 Päivi Paukku & Jenni Laine Centre for Language and Communication Studies
 Information on Finnish Language Courses Spring Semester 2018 Päivi Paukku & Jenni Laine 4.1.2018 Centre for Language and Communication Studies Puhutko suomea? -Hei! -Hei hei! -Moi! -Moi moi! -Terve! -Terve
Information on Finnish Language Courses Spring Semester 2018 Päivi Paukku & Jenni Laine 4.1.2018 Centre for Language and Communication Studies Puhutko suomea? -Hei! -Hei hei! -Moi! -Moi moi! -Terve! -Terve
Siirtymä maisteriohjelmiin tekniikan korkeakoulujen välillä Transfer to MSc programmes between engineering schools
 Siirtymä maisteriohjelmiin tekniikan korkeakoulujen välillä Transfer to MSc programmes between engineering schools Akateemisten asioiden komitea Academic Affairs Committee 11 October 2016 Eija Zitting
Siirtymä maisteriohjelmiin tekniikan korkeakoulujen välillä Transfer to MSc programmes between engineering schools Akateemisten asioiden komitea Academic Affairs Committee 11 October 2016 Eija Zitting
Rakennukset Varjostus "real case" h/a 0,5 1,5
 Tuulivoimala Rakennukset Asuinrakennus Liikerak. tai Julkinen rak. Lomarakennus Teollinen rakennus Kirkollinen rakennus Varjostus "real case" h/a 1 h/a 8 h/a 20 h/a 1 2 3 5 8 4 6 7 9 10 0 0,5 1 1,5 2 km
Tuulivoimala Rakennukset Asuinrakennus Liikerak. tai Julkinen rak. Lomarakennus Teollinen rakennus Kirkollinen rakennus Varjostus "real case" h/a 1 h/a 8 h/a 20 h/a 1 2 3 5 8 4 6 7 9 10 0 0,5 1 1,5 2 km
I. AES Rijndael. Rijndael - Internal Structure
 I. AES Rndael NOKIA T-79.53 Additional material Oct 3/KN Rndael - Internal Structure Rndael is an iterated block cipher with variable length block and variable key size. The number of rounds is defined
I. AES Rndael NOKIA T-79.53 Additional material Oct 3/KN Rndael - Internal Structure Rndael is an iterated block cipher with variable length block and variable key size. The number of rounds is defined
Käyttöliittymät II. Käyttöliittymät I Kertaus peruskurssilta. Keskeisin kälikurssilla opittu asia?
 Käyttöliittymät II Sari A. Laakso Käyttöliittymät I Kertaus peruskurssilta Keskeisin kälikurssilla opittu asia? 1 Käyttöliittymät II Kurssin sisältö Käli I Käyttötilanteita Käli II Käyttötilanteet selvitetään
Käyttöliittymät II Sari A. Laakso Käyttöliittymät I Kertaus peruskurssilta Keskeisin kälikurssilla opittu asia? 1 Käyttöliittymät II Kurssin sisältö Käli I Käyttötilanteita Käli II Käyttötilanteet selvitetään
FIS IMATRAN KYLPYLÄHIIHDOT Team captains meeting
 FIS IMATRAN KYLPYLÄHIIHDOT 8.-9.12.2018 Team captains meeting 8.12.2018 Agenda 1 Opening of the meeting 2 Presence 3 Organizer s personell 4 Jury 5 Weather forecast 6 Composition of competitors startlists
FIS IMATRAN KYLPYLÄHIIHDOT 8.-9.12.2018 Team captains meeting 8.12.2018 Agenda 1 Opening of the meeting 2 Presence 3 Organizer s personell 4 Jury 5 Weather forecast 6 Composition of competitors startlists
Research plan for masters thesis in forest sciences. The PELLETime 2009 Symposium Mervi Juntunen
 Modelling tree and stand characteristics and estimating biomass removals and harvesting costs of lodgepole pine (Pinus contorta) plantations in Iceland Research plan for masters thesis in forest sciences
Modelling tree and stand characteristics and estimating biomass removals and harvesting costs of lodgepole pine (Pinus contorta) plantations in Iceland Research plan for masters thesis in forest sciences
3 9-VUOTIAIDEN LASTEN SUORIUTUMINEN BOSTONIN NIMENTÄTESTISTÄ
 Puhe ja kieli, 27:4, 141 147 (2007) 3 9-VUOTIAIDEN LASTEN SUORIUTUMINEN BOSTONIN NIMENTÄTESTISTÄ Soile Loukusa, Oulun yliopisto, suomen kielen, informaatiotutkimuksen ja logopedian laitos & University
Puhe ja kieli, 27:4, 141 147 (2007) 3 9-VUOTIAIDEN LASTEN SUORIUTUMINEN BOSTONIN NIMENTÄTESTISTÄ Soile Loukusa, Oulun yliopisto, suomen kielen, informaatiotutkimuksen ja logopedian laitos & University
The Viking Battle - Part Version: Finnish
 The Viking Battle - Part 1 015 Version: Finnish Tehtävä 1 Olkoon kokonaisluku, ja olkoon A n joukko A n = { n k k Z, 0 k < n}. Selvitä suurin kokonaisluku M n, jota ei voi kirjoittaa yhden tai useamman
The Viking Battle - Part 1 015 Version: Finnish Tehtävä 1 Olkoon kokonaisluku, ja olkoon A n joukko A n = { n k k Z, 0 k < n}. Selvitä suurin kokonaisluku M n, jota ei voi kirjoittaa yhden tai useamman
MUSEOT KULTTUURIPALVELUINA
 Elina Arola MUSEOT KULTTUURIPALVELUINA Tutkimuskohteena Mikkelin museot Opinnäytetyö Kulttuuripalvelujen koulutusohjelma Marraskuu 2005 KUVAILULEHTI Opinnäytetyön päivämäärä 25.11.2005 Tekijä(t) Elina
Elina Arola MUSEOT KULTTUURIPALVELUINA Tutkimuskohteena Mikkelin museot Opinnäytetyö Kulttuuripalvelujen koulutusohjelma Marraskuu 2005 KUVAILULEHTI Opinnäytetyön päivämäärä 25.11.2005 Tekijä(t) Elina